
Войти
 Представление трехмерной структуры белка миоглобин показаны бирюзовые α-спирали. Этот белок был первым, чья структура была определена с помощью рентгеновской кристаллографии. Ближе к правому центру среди спиралей простетическая группа, называемая гемовой группой (показана серым цветом) со связанной молекулой кислорода (красный).
Представление трехмерной структуры белка миоглобин показаны бирюзовые α-спирали. Этот белок был первым, чья структура была определена с помощью рентгеновской кристаллографии. Ближе к правому центру среди спиралей простетическая группа, называемая гемовой группой (показана серым цветом) со связанной молекулой кислорода (красный). Белки являются большие биомолекулы или макромолекулы, состоящие из одной или нескольких длинных цепей аминокислотных остатков. Белки выполняют широкий спектр функций внутри организмов, в том числе катализировать метаболические реакции, репликацию ДНК, отвечать на стимулы, обеспечивая структуру клеткам, и организмы, и , транспортирующие молекулы из одного места в другое. Белки отличаются друг от друга в первую очередь аминокислотной последовательностью, которая определяется нуклеотидной последовательностью их генов, и которая обычно приводит к сворачиванию белка в конкретная трехмерная структура, определяющая его активность.
Линейная цепь аминокислотных остатков называется полипептидом. Белок содержит по крайней мере один длинный полипептид. Короткие полипептиды, содержащие менее 20–30 остатков, редко считаются белками и обычно называются пептидами или иногда олигопептидами. Отдельные аминокислотные остатки связаны вместе пептидными связями и соседними аминокислотными остатками. Последовательность аминокислотных остатков в белке определяется последовательностью гена , которая кодируется генетическим кодом. В общем, генетический код определяет 20 стандартных аминокислот; но у некоторых организмов генетический код может включать селеноцистеин и - в некоторых архей - пирролизин. Вскоре после или даже во время синтеза остатки в белке часто химически модифицируются с помощью посттрансляционной модификации, которая изменяет физические и химические свойства, фолдинг, стабильность, активность и, в конечном итоге, функцию белков.. К некоторым белкам присоединены непептидные группы, которые можно назвать простетическими группами или кофакторами. Белки также могут работать вместе для достижения определенной функции, и они часто объединяются, образуя стабильные белковые комплексы.
После образования белки существуют только в течение определенного периода, а затем разлагаются и повторно используются клеточный аппарат в процессе белкового обмена. Продолжительность жизни белка измеряется с помощью его периода полужизни и охватывает широкий диапазон. Они могут существовать в течение нескольких минут или лет, а средняя продолжительность жизни в клетках млекопитающих составляет 1-2 дня. Аномальные или неправильно свернутые белки разлагаются быстрее либо из-за того, что они нацелены на разрушение, либо из-за нестабильности.
Подобно другим биологическим макромолекулам, таким как полисахариды и нуклеиновые кислоты, белки являются важными частями организмов и участвуют практически во всех процессах внутри клеток. Многие белки являются ферментами, которые катализируют биохимические реакции и жизненно важны для метаболизма. Белки также выполняют структурные или механические функции, такие как актин и миозин в мышцах и белки в цитоскелете, которые образуют систему каркаса, который поддерживает форму ячейки. Другие белки важны в передаче сигналов, иммунных ответах, адгезии клеток и клеточном цикле. У животных белки необходимы в диете для обеспечения незаменимыми аминокислотами, которые невозможно синтезировать. Пищеварение расщепляет белки для использования в метаболизме.
Белки могут быть очищены от других клеточных компонентов с использованием различных методов, таких как ультрацентрифугирование, осаждение, электрофорез и хроматография ; Появление генной инженерии сделало возможным ряд методов, облегчающих очистку. Методы, обычно используемые для изучения структуры и функции белков, включают иммуногистохимию, сайт-направленный мутагенез, рентгеновскую кристаллографию, ядерный магнитный резонанс и масс-спектрометрия.
Белки были признаны отдельным классом биологических молекул в восемнадцатом веке Антуаном Фуркроем и другими, отличавшимися способностью молекул к коагулируйте или флокулируйте под действием тепла или кислоты. Среди отмеченных примеров в то время были альбумин из яичных белков, сывороточный альбумин крови, фибрин и пшеничный глютен.
. Белки были впервые описаны Голландский химик Герардус Йоханнес Малдер и назван шведским химиком Йенсом Якобом Берцелиусом в 1838 году. Малдер провел элементный анализ обычных белков и обнаружил, что почти все белки имеют та же эмпирическая формула, C 400 H 620 N 100 O 120 P1S1. Он пришел к ошибочному выводу, что они могут состоять из одного типа (очень больших) молекул. Термин «белок» для описания этих молекул был предложен сотрудником Малдера Берцелиусом; белок происходит от греческого слова πρώτειος (proteios), что означает «первичный», «впереди» или «стоящий впереди», + -in. Малдер продолжил идентифицировать продукты деградации белка, такие как аминокислота лейцин, для которой он обнаружил (почти правильную) молекулярную массу 131 Da. До слова «белок» использовались другие названия, такие как «альбумины» или «белковые материалы» (Eiweisskörper, на немецком языке).
Ранние ученые-диетологи, такие как немецкий Карл фон Войт, считали этот белок был наиболее важным питательным веществом для поддержания структуры тела, потому что обычно считалось, что «плоть рождает плоть». Карл Генрих Риттхаузен расширил известные формы белка с идентификацией глутаминовой кислоты. На сельскохозяйственной экспериментальной станции Коннектикута подробный обзор растительных белков был составлен Томасом Берром Осборном. Работая с Лафайетом Менделем и применяя закон минимума в кормлении лабораторных крыс, были установлены питательные незаменимые аминокислоты. Работа была продолжена и сообщена Уильямом Каммингом Роуз. Понимание белков как полипептидов появилось благодаря работе Франца Хофмайстера и Германа Эмиля Фишера в 1902 году. Центральная роль белков как ферментов в живых организмах не было полностью изучено до 1926 года, когда Джеймс Б. Самнер показал, что фермент уреаза на самом деле является белком.
Сложность очистки белков в больших количествах делали их очень трудными для изучения первыми биохимиками белков. Следовательно, ранние исследования были сосредоточены на белках, которые можно было очищать в больших количествах, например, белков крови, яичного белка, различных токсинов и пищеварительных / метаболических ферментов, полученных на бойнях. В 1950-х годах Armor Hot Dog Co. очистила 1 кг чистой бычьей панкреатической рибонуклеазы A и сделала ее свободно доступной для ученых; этот жест помог рибонуклеазе А стать основной целью биохимических исследований в последующие десятилетия.
 Джон Кендрю с моделью миоглобина в разработке
Джон Кендрю с моделью миоглобина в разработке Лайнусу Полингу приписывают успешное предсказание регулярного белка вторичные структуры, основанные на водородной связи, идея, впервые выдвинутая Уильямом Эстбери в 1933 году. Более поздняя работа Уолтера Каузмана по денатурации, частично основанный на предыдущих исследованиях Кай Линдерстрём-Ланг, внес вклад в понимание сворачивания белка и структуры, опосредованной гидрофобными взаимодействиями.
Первый белок, который будет секвенированный был инсулином, Фредериком Сэнгером в 1949 году. Сэнгер правильно определил аминокислотную последовательность инсулина, тем самым убедительно продемонстрировав, что белки состоят из линейных полимеров аминокислотных остатков. кислоты, а не с разветвленными цепями, коллоиды или циклолы. За это достижение он получил Нобелевскую премию в 1958 году.
Первыми белковыми структурами, которые нужно было решить, были гемоглобин и миоглобин на <256.>Макс Перуц и сэр Джон Каудери Кендрю, соответственно, в 1958 году. По состоянию на 2017 год в Protein Data Bank содержится более 126060 структур белков с атомным разрешением. В последнее время криоэлектронная микроскопия больших макромолекулярных ансамблей и расчетное прогнозирование структуры белка малых белковых доменов являются двумя методами, приближающимися к атомное разрешение.
 Химическая структура пептидной связи (внизу) и трехмерная структура пептидной связи между аланином и соседней аминокислотой (вверху / вставка). Сама связь состоит из элементов CHON.
Химическая структура пептидной связи (внизу) и трехмерная структура пептидной связи между аланином и соседней аминокислотой (вверху / вставка). Сама связь состоит из элементов CHON. 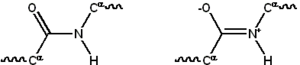 Резонансные структуры пептидной связи, которая связывает отдельные аминокислоты с образованием белка полимера
Резонансные структуры пептидной связи, которая связывает отдельные аминокислоты с образованием белка полимера Большинство белков состоит из линейных полимеров, построенных из ряда до 20 различных L-α- аминокислот. Все протеиногенные аминокислоты обладают общими структурными особенностями, включая α-углерод, к которому относится амино группа, карбоксильная группа и переменная боковая цепь связана. Только пролин отличается от этой базовой структуры, поскольку он содержит кольцо, необычное для N-концевой аминогруппы, которое заставляет амидный фрагмент CO – NH принимать фиксированную конформацию. Боковые цепи стандартных аминокислот, подробно описанные в списке стандартных аминокислот, имеют большое разнообразие химических структур и свойств; это комбинированное действие всех боковых цепей аминокислот в белке, которое в конечном итоге определяет его трехмерную структуру и его химическую реактивность. Аминокислоты в полипептидной цепи связаны пептидными связями. Однажды связанная в белковой цепи отдельная аминокислота называется остатком, а связанный ряд атомов углерода, азота и кислорода известен как основная цепь или белковый каркас.
Пептидная связь имеет два резонансные формы, которые вносят некоторый характер двойной связи и препятствуют вращению вокруг своей оси, так что альфа-атомы углерода являются примерно копланарными. Два других двугранных угла в пептидной связи определяют локальную форму, которую принимает остов белка. Конец со свободной аминогруппой известен как N-конец или амино-конец, тогда как конец белка со свободной карбоксильной группой известен как С-конец или карбокси. конец (последовательность белка записывается от N-конца к C-концу слева направо).
Слова белок, полипептид и пептид несколько неоднозначны и могут перекрываться по значению. Белок обычно используется для обозначения полной биологической молекулы в стабильной конформации, тогда как пептид обычно зарезервирован для олигомеров с короткими аминокислотами, часто не имеющими стабильной трехмерной структуры. Но граница между ними четко не определена и обычно проходит около 20–30 остатков. Полипептид может относиться к любой одиночной линейной цепи аминокислот, обычно независимо от длины, но часто подразумевает отсутствие определенной конформации.
Белки могут взаимодействовать со многими типами молекул, включая с другими белками, с липидами, с углеводами и с ДНК.
Было подсчитано, что Бактерии среднего размера содержат около 2 миллионов белков на клетку (например, E. coli и Staphylococcus aureus ). Более мелкие бактерии, такие как Mycoplasma или спирохеты, содержат меньше молекул, от 50 000 до 1 миллиона. Напротив, эукариотические клетки больше и, следовательно, содержат гораздо больше белка. Например, дрожжевые клетки, по оценкам, содержат около 50 миллионов белков, а человеческие клетки - порядка от 1 до 3 миллиардов. Концентрация отдельных белковых копий колеблется от нескольких молекул на клетку до 20 миллионов. Не все гены, кодирующие белки, экспрессируются в большинстве клеток, и их количество зависит, например, от типа клетки и внешних стимулов. Например, из 20 000 или около того белков, кодируемых геномом человека, только 6000 выявляются в лимфобластоидных клетках. Более того, количество белков, кодируемых геномом, хорошо коррелирует со сложностью организма. У эукариот - 15000, бактерий - 3200, архей - 2400, а вирусы - в среднем 42 белка, закодированных в их соответствующих геномах.
 Последовательность ДНК гена кодирует аминокислотную последовательность белка
Последовательность ДНК гена кодирует аминокислотную последовательность белка Белки собираются из аминокислотной последовательности. кислоты с использованием информации, закодированной в генах. Каждый белок имеет собственную уникальную аминокислотную последовательность, которая определяется последовательностью нуклеотидов гена, кодирующего этот белок. Генетический код представляет собой набор из трех нуклеотидных наборов, называемых кодонами, и каждая комбинация из трех нуклеотидов обозначает аминокислоту, например, AUG (аденин - урацил - гуанин ) - это код для метионина. Поскольку ДНК содержит четыре нуклеотида, общее количество возможных кодонов составляет 64; следовательно, в генетическом коде присутствует некоторая избыточность, причем некоторые аминокислоты определяются более чем одним кодоном. Гены, кодируемые в ДНК, сначала транскрибируются в пре- информационную РНК (мРНК) такими белками, как РНК-полимераза. Большинство организмов затем обрабатывают пре-мРНК (также известную как первичный транскрипт) с использованием различных форм посттранскрипционной модификации с образованием зрелой мРНК, которая затем используется в качестве матрицы для синтеза белка рибосома. В прокариотах мРНК может либо использоваться, как только она продуцируется, либо связываться с рибосомой после того, как она отошла от нуклеоида. Напротив, эукариоты производят мРНК в ядре клетки, а затем перемещают ее через ядерную мембрану в цитоплазму, где затем происходит синтез белка . Скорость синтеза белка у прокариот выше, чем у эукариот, и может достигать 20 аминокислот в секунду.
Процесс синтеза белка из матрицы мРНК известен как трансляция. МРНК загружается на рибосому и считывается с трех нуклеотидов за раз путем сопоставления каждого кодона с его парой оснований антикодоном, расположенным на молекуле транспортной РНК, которая несет аминокислоту, соответствующую распознаваемому кодону. Фермент аминоацил тРНК синтетаза «заряжает» молекулы тРНК правильными аминокислотами. Растущий полипептид часто называют растущей цепью. Белки всегда биосинтезируются от N-конца до C-конца.
Размер синтезированного белка можно измерить по количеству содержащихся в нем аминокислот и по его общей молекулярной массе., который обычно указывается в единицах дальтон (синоним единиц атомной массы ) или производной единице килодальтона (кДа). Средний размер белка увеличивается от архей к бактериям и эукариотам (283, 311, 438 остатков и 31, 34, 49 кДа соответственно) из-за большего числа белковых доменов, составляющих белки у высших организмов. Например, дрожжевые белки в среднем состоят из 466 аминокислот в длину и 53 кДа по массе. Самыми крупными известными белками являются тайтины, компонент мышцы саркомера, с молекулярной массой почти 3000 кДа и общей длиной почти 27000 аминокислот..
Короткие белки также могут быть синтезированы химическим путем с помощью семейства методов, известных как пептидный синтез, которые основаны на методах органического синтеза такие как химическое лигирование для получения пептидов с высоким выходом. Химический синтез позволяет вводить неприродные аминокислоты в полипептидные цепи, например прикреплять флуоресцентные зонды к боковым цепям аминокислот. Эти методы полезны в лабораторной биохимии и клеточной биологии, хотя, как правило, не для коммерческого применения. Химический синтез неэффективен для полипептидов, длина которых превышает примерно 300 аминокислот, и синтезированные белки не могут легко принять свою природную третичную структуру. Большинство методов химического синтеза осуществляются от С-конца к N-концу, в противоположность биологической реакции.
 Кристаллическая структура шаперонина, огромного белкового комплекса. Выделена одна белковая субъединица. Шаперонины способствуют сворачиванию белка.
Кристаллическая структура шаперонина, огромного белкового комплекса. Выделена одна белковая субъединица. Шаперонины способствуют сворачиванию белка. 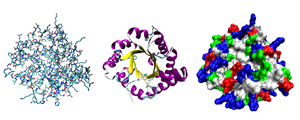 Три возможных представления трехмерной структуры белка триозофосфат-изомеразы. Слева : представление всего атома, окрашенное по типу атома. В середине: Упрощенное представление, иллюстрирующее конформацию позвоночника, окрашенную вторичной структурой. Справа : доступное для растворителя изображение поверхности, окрашенное по типу остатка (кислотные остатки - красный, основные остатки - синий, полярные остатки - зеленый, неполярные остатки - белые).
Три возможных представления трехмерной структуры белка триозофосфат-изомеразы. Слева : представление всего атома, окрашенное по типу атома. В середине: Упрощенное представление, иллюстрирующее конформацию позвоночника, окрашенную вторичной структурой. Справа : доступное для растворителя изображение поверхности, окрашенное по типу остатка (кислотные остатки - красный, основные остатки - синий, полярные остатки - зеленый, неполярные остатки - белые). Большинство белков складываются в уникальное трехмерное изображение конструкции. Форма, в которую естественным образом складывается белок, известна как его нативная конформация. Хотя многие белки могут сворачиваться без посторонней помощи, просто благодаря химическим свойствам их аминокислот, другим требуется помощь молекулярных шаперонов, чтобы укладываться в их нативное состояние. Биохимики часто ссылаются на четыре отдельных аспекта структуры белка:
Белки не являются полностью жесткими молекулами. В дополнение к этим уровням структуры белки могут перемещаться между несколькими родственными структурами, пока они выполняют свои функции. В контексте этих функциональных перестроек эти третичные или четвертичные структуры обычно упоминаются как «конформации », а переходы между ними называются конформационными изменениями. Такие изменения часто индуцируются связыванием молекулы субстрата с активным сайтом фермента или физической областью белка, которая участвует в химическом катализе. В растворе белки также претерпевают изменения в структуре из-за тепловых колебаний и столкновений с другими молекулами.
 Молекулярная поверхность нескольких белков, показывающая их сравнительные размеры. Слева направо: иммуноглобулин G (IgG, антитело ), гемоглобин, инсулин (гормон), аденилаткиназа (фермент) и глутамин синтетаза (фермент).
Молекулярная поверхность нескольких белков, показывающая их сравнительные размеры. Слева направо: иммуноглобулин G (IgG, антитело ), гемоглобин, инсулин (гормон), аденилаткиназа (фермент) и глутамин синтетаза (фермент). Белки можно неофициально разделить на три основных класса, которые коррелируют с типичными третичными структурами: глобулярные белки, волокнистые белки и мембранные белки. Почти все глобулярные белки растворимы, и многие из них являются ферментами. Волокнистые белки часто являются структурными, такими как коллаген, главный компонент соединительной ткани, или кератин, белковый компонент волос и ногтей. Мембранные белки часто служат рецепторами или обеспечивают каналы для полярных или заряженных молекул, проходящих через клеточную мембрану.
. Особый случай внутримолекулярных водородных связей внутри белков, плохо защищенных от воздействия воды и, следовательно, способствующих их собственная дегидратация, называются дегидронами.
Многие белки состоят из нескольких белковых доменов, то есть сегментов белка, которые складываются в отдельные структурные единицы. Домены обычно также имеют определенные функции, такие как ферментативная активность (например, киназа ), или они служат в качестве связывающих модулей (например, домен SH3 связывается с последовательностями, богатыми пролином в других белках).
Короткие аминокислотные последовательности в белках часто действуют как сайты узнавания для других белков. Например, домены SH3 обычно связываются с короткими мотивами PxxP (т. Е. 2 пролина [P], разделенных двумя неуказанными аминокислотами [x], хотя окружающие аминокислоты могут определять точную специфичность связывания). Многие такие мотивы собраны в базе данных Eukaryotic Linear Motif (ELM).
Белки - это главные действующие лица клетки, которые, как говорят, выполняют функции, определенные информацией, закодированной в генах. За исключением некоторых типов РНК, большинство других биологических молекул являются относительно инертными элементами, на которые действуют белки. Белки составляют половину сухой массы клетки Escherichia coli, тогда как другие макромолекулы, такие как ДНК и РНК, составляют только 3% и 20% соответственно. Набор белков, экспрессируемых в конкретной клетке или типе клеток, известен как его протеом.
 . Фермент гексокиназа показан в виде обычной молекулярной модели типа шарик и палочка. В правом верхнем углу в масштабе показаны два его субстрата, АТФ и глюкоза.
. Фермент гексокиназа показан в виде обычной молекулярной модели типа шарик и палочка. В правом верхнем углу в масштабе показаны два его субстрата, АТФ и глюкоза.. Главной характеристикой белков, которая также обеспечивает их разнообразный набор функций, является их способность связывать другие молекулы. конкретно и плотно. Область белка, отвечающая за связывание другой молекулы, известна как сайт связывания и часто представляет собой углубление или «карман» на поверхности молекулы. Эта связывающая способность опосредована третичной структурой белка, которая определяет карман сайта связывания, и химическими свойствами боковых цепей окружающих аминокислот. Связывание с белками может быть чрезвычайно прочным и специфичным; например, белок ингибитор рибонуклеазы связывается с человеческим ангиогенином с субфемтомолярной константой диссоциации (<10 M) but does not bind at all to its amphibian homolog онконаза (>1 M). Чрезвычайно незначительные химические изменения, такие как добавление одной метильной группы к связывающему партнеру, иногда могут быть достаточными для почти полного устранения связывания; например, аминоацил тРНК-синтетаза, специфичная для аминокислоты валин различает очень похожую боковую цепь аминокислоты изолейцин.
. Белки могут связываться с другими белками, а также с низкомолекулярными субстратами. Когда белки специфически связываются с другими копиями той же молекулы, они могут олигомеризоваться с образованием фибрилл; этот процесс часто происходит в структурных белках, которые состоят из глобулярных мономеров, которые самоассоциируются с образованием жестких волокон. Белковые взаимодействия также регулируют ферментативную активность, контролируют прохождение клеточного цикла и позволяют собирать большие протеиновые в комплексах, которые выполняют множество тесно связанных реакций с общей биологической функцией. Белки также могут связываться с клеточными мембранами или даже интегрироваться в них. Способность партнеров по связыванию вызывать конформационные изменения в белках позволяет создавать чрезвычайно сложные сигнальные сети. Поскольку взаимодействия между белками обратимы и в значительной степени зависят от доступности различных групп белков-партнеров для формирования агрегатов, способных выполнять дискретные наборы функций, изучение взаимодействий между конкретными белками является ключом к пониманию важных аспектов клеточной функции. и, в конечном итоге, свойства, которые различают определенные типы клеток.
Самая известная роль белков в клетке - это как ферменты, которые катализируют химические реакции. Ферменты обычно очень специфичны и ускоряют только одну или несколько химических реакций. Ферменты осуществляют большинство реакций, участвующих в метаболизме, а также манипулируют ДНК в таких процессах, как репликация ДНК, репарация ДНК и транскрипция. Некоторые ферменты воздействуют на другие белки, добавляя или удаляя химические группы в процессе, известном как посттрансляционная модификация. Известно, что ферменты катализируют около 4000 реакций. Ускорение скорости, обеспечиваемое ферментативным катализом, часто бывает огромным - до 10-кратного увеличения скорости по сравнению с некаталитической реакцией в случае оротатдекарбоксилазы (78 миллионов лет без фермента, 18 миллисекунд с ферментом).
Молекулы, связанные ферментами и действующие на них, называются субстратами. Хотя ферменты могут состоять из сотен аминокислот, обычно только небольшая часть остатков контактирует с субстратом, а еще меньшая часть - в среднем от трех до четырех остатков - непосредственно участвует в катализе. Область фермента, которая связывает субстрат и содержит каталитические остатки, известна как активный центр.
Диригентные белки являются членами класса белков, которые определяют стереохимию соединение, синтезируемое другими ферментами.
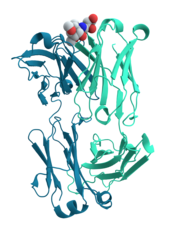 Ленточная диаграмма мышиного антитела против холеры, которое связывает углеводный антиген
Ленточная диаграмма мышиного антитела против холеры, которое связывает углеводный антиген Многие белки участвуют в процессе передачи сигналов клеток и передачи сигналов. Некоторые белки, такие как инсулин, являются внеклеточными белками, которые передают сигнал от клетки, в которой они были синтезированы, другим клеткам в отдаленных тканях. Другими являются мембранные белки, которые действуют как рецепторы, основная функция которых заключается в связывании сигнальной молекулы и индукции биохимического ответа в клетке. Многие рецепторы имеют сайт связывания, расположенный на поверхности клетки, и эффекторный домен внутри клетки, который может обладать ферментативной активностью или может подвергаться конформационному изменению, обнаруживаемому другими белками внутри клетки.
Антитела представляют собой белковые компоненты адаптивной иммунной системы, основная функция которых заключается в связывании антигенов или чужеродных веществ в организме и нацеливании на них для разрушения. Антитела могут секретироваться во внеклеточную среду или заякорены в мембранах специализированных В-клеток, известных как плазматические клетки. В то время как ферменты ограничены в их аффинности связывания со своими субстратами из-за необходимости проведения реакции, антитела не имеют таких ограничений. Аффинность связывания антитела с его мишенью чрезвычайно высока.
Многие белки-транспортеры лигандов связывают определенные небольшие биомолекулы и транспортируют их в другие места в теле многоклеточного организма. Эти белки должны обладать высокой аффинностью связывания, когда их лиганд присутствует в высоких концентрациях, но также должны высвобождать лиганд, когда он присутствует в низких концентрациях в тканях-мишенях. Каноническим примером лиганд-связывающего белка является гемоглобин, который транспортирует кислород из легких в другие органы и ткани у всех позвоночных и имеет близких гомологов в каждом биологическом царстве. Лектины представляют собой сахаросвязывающие белки, которые высокоспецифичны для своих сахарных фрагментов. Лектины обычно играют роль в явлениях биологического распознавания с участием клеток и белков. Рецепторы и гормоны представляют собой высокоспецифичные связывающие белки.
Трансмембранные белки могут также служить белками-переносчиками лигандов, которые изменяют проницаемость клеточной мембраны для малых молекул и ионов. Сама по себе мембрана имеет гидрофобное ядро, через которое полярные или заряженные молекулы не могут диффундировать. Мембранные белки содержат внутренние каналы, которые позволяют таким молекулам входить и выходить из клетки. Многие белки ионного канала специализируются на выборе только определенного иона; например, каналы калия и натрия часто различают только один из двух ионов.
.
Структурные белки придают жесткость и жесткость жидкостям биологические компоненты. Большинство структурных белков - это волокнистые белки ; например, коллаген и эластин являются критическими компонентами соединительной ткани, например хрящ, и обнаружен кератин в твердых или нитчатых структурах, таких как волосы, ногти, перья, копыта и некоторые панцири животных. Некоторые глобулярные белки также могут выполнять структурные функции, например, актин и тубулин глобулярны и растворимы как мономеры, но полимеризуются с образованием длинные жесткие волокна, составляющие цитоскелет, что позволяет клетке сохранять свою форму и размер.
Другими белками, которые выполняют структурные функции, являются моторные белки, такие как миозин, кинезин и динеин, которые являются способен генерировать механические силы. Эти белки имеют решающее значение для клеточной подвижности одноклеточных организмов и сперматозоидов многих многоклеточных организмов, которые воспроизводятся половым путем. Они также генерируют силы, возникающие при сокращении мышц, и играют важную роль во внутриклеточном транспорте.
Активность и структуру белков можно исследовать in vitro, in vivo и in silico. In vitro исследования очищенных белков в контролируемой среде полезны для изучения того, как белок выполняет свою функцию: например, исследования кинетики фермента исследуют химический механизм каталитическая активность фермента и его относительное сродство к различным возможным молекулам субстрата. Напротив, эксперименты in vivo могут предоставить информацию о физиологической роли белка в контексте клетки или даже всего организма. In silico исследования используют вычислительные методы для изучения белков.
Для выполнения анализа in vitro белок должен быть очищен от других клеточных компонентов. Этот процесс обычно начинается с лизиса клеток, при котором мембрана клетки разрушается и ее внутреннее содержимое высвобождается в раствор, известный как неочищенный лизат. Полученную смесь можно очистить с помощью ультрацентрифугирования, которое фракционирует различные клеточные компоненты на фракции, содержащие растворимые белки; мембранные липиды и белки; клеточные органеллы и нуклеиновые кислоты. Осаждение методом, известным как высаливание, может концентрировать белки из этого лизата. Затем используются различные типы хроматографии для выделения интересующего белка или белков на основе таких свойств, как молекулярная масса, общий заряд и сродство связывания. Уровень очистки можно контролировать с помощью различных типов гель-электрофореза, если известны желаемая молекулярная масса белка и изоэлектрическая точка, с помощью спектроскопии, если белок имеет различимый спектроскопическими признаками или с помощью ферментных анализов, если белок обладает ферментативной активностью. Кроме того, белки могут быть выделены в соответствии с их зарядом с помощью электрофокусировки.
. Для природных белков может потребоваться серия стадий очистки, чтобы получить белок, достаточно чистый для лабораторных применений. Чтобы упростить этот процесс, часто используется генная инженерия для добавления химических свойств к белкам, которые упрощают их очистку, не влияя на их структуру или активность. Здесь «метка», состоящая из конкретной аминокислотной последовательности, часто из серии остатков гистидина («His-метка »), прикреплена к одному концу белка. В результате, когда лизат пропускают через хроматографическую колонку, содержащую никель, остатки гистидина связывают никель и присоединяются к колонке, в то время как немаркированные компоненты лизата проходят беспрепятственно. Был разработан ряд различных тегов, чтобы помочь исследователям очистить определенные белки от сложных смесей.
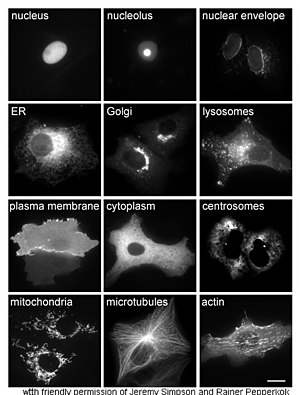 Белки в различных клеточных компартментах и структурах, помеченных зеленым флуоресцентным белком (здесь белый)
Белки в различных клеточных компартментах и структурах, помеченных зеленым флуоресцентным белком (здесь белый) Изучение белков in vivo часто связано с синтезом и локализацией белка внутри клетки. Хотя многие внутриклеточные белки синтезируются в цитоплазме и мем Связанные с браной или секретируемые белки в эндоплазматическом ретикулуме, особенности того, как белки нацелены на конкретные органеллы или клеточные структуры, часто неясны. Полезный метод оценки клеточной локализации использует генную инженерию для экспрессии в клетке слитого белка или химеры, состоящего из интересующего природного белка, связанного с «репортером "такие как зеленый флуоресцентный белок (GFP). Положение слитого белка в клетке можно четко и эффективно визуализировать с помощью микроскопии, как показано на рисунке напротив.
Другие методы выяснения клеточного местоположения белков требуют использования известных компартментных маркеров для таких областей, как ER, Гольджи, лизосомы или вакуоли, митохондрии, хлоропласты, плазматическая мембрана и т. Д. С использованием флуоресцентного меченые версии этих маркеров или антител к известным маркерам, становится намного проще определить локализацию интересующего белка. Например, непрямая иммунофлуоресценция позволит колокализацию флуоресценции и демонстрацию местоположения. Флуоресцентные красители используются для маркировки клеточных компартментов с аналогичной целью.
Существуют и другие возможности. Например, в иммуногистохимии обычно используются антитела к одному или нескольким интересующим белкам, которые конъюгированы с ферментами, дающими люминесцентные или хромогенные сигналы, которые можно сравнивать между образцами, что позволяет получить информацию о локализации. Другой применимый метод - это совместное фракционирование в градиентах сахарозы (или другого материала) с использованием изопикнического центрифугирования. Хотя этот метод не доказывает совместную локализацию компартмента известной плотности и интересующего белка, он увеличивает вероятность и более поддается крупномасштабным исследованиям.
Наконец, золотым стандартом клеточной локализации является иммуноэлектронная микроскопия. В этом методе также используются антитела к интересующему белку наряду с классическими методами электронной микроскопии. Образец готовят для обычного исследования под электронным микроскопом, а затем обрабатывают антителом к интересующему белку, который конъюгирован с чрезвычайно электроплотным материалом, обычно золотом. Это позволяет локализовать как детали ультраструктуры, так и интересующий белок.
С помощью другого приложения генной инженерии, известного как сайт-направленный мутагенез, исследователи могут изменять последовательность белка и, следовательно, его структура, клеточная локализация и восприимчивость к регуляции. Этот метод позволяет даже включать неприродные аминокислоты в белки с использованием модифицированных тРНК и может позволить рациональное проектирование новых белков с новыми свойствами.
Общий набор белков, присутствующих в определенный момент в клетке или типе клеток, известен как его протеом, и изучение таких крупномасштабных наборов данных определяет область протеомики, названную аналогия со смежной областью геномики. Ключевые экспериментальные методы в протеомике включают 2D-электрофорез, который позволяет разделить многие белки, масс-спектрометрию, которая позволяет быстро идентифицировать белки с высокой пропускной способностью и секвенировать пептиды (чаще всего после расщепление в геле ), белковые микроматрицы, которые позволяют определять относительные уровни различных белков, присутствующих в клетке, и двухгибридный скрининг, который позволяет систематически исследовать белок-белковые взаимодействия. Полный набор биологически возможных таких взаимодействий известен как интерактом. Систематическая попытка определить структуры белков, представляющих все возможные складки, известна как структурная геномика.
Для анализа структуры, функции и эволюции белков был разработан широкий спектр вычислительных методов..
Разработка таких инструментов была обусловлена большим объемом геномных и протеомных данных, доступных для различных организмов, включая геном человека. Экспериментально изучить все белки просто невозможно, поэтому только некоторые из них подвергаются лабораторным экспериментам, в то время как вычислительные инструменты используются для экстраполяции на похожие белки. Такие гомологичные белки можно эффективно идентифицировать в отдаленно родственных организмах с помощью выравнивания последовательностей. Геном и последовательности генов можно искать с помощью различных инструментов на предмет определенных свойств. Инструменты профилирования последовательностей могут находить сайты рестрикционных ферментов, открытые рамки считывания в нуклеотидных последовательностях и предсказывать вторичные структуры. Филогенетические деревья могут быть построены, а эволюционные гипотезы могут быть разработаны с использованием специального программного обеспечения, такого как ClustalW, в отношении происхождения современных организмов и генов, которые они экспрессируют. Область биоинформатики сейчас незаменима для анализа генов и белков.
Обнаружение третичной структуры белка или четвертичной структуры его комплексов может дать важные подсказки о том, как белок выполняет свою функцию и как на нее можно повлиять, т.е. дизайн препарата. Поскольку белки слишком малы, чтобы их можно было увидеть под световым микроскопом, для определения их структуры необходимо использовать другие методы. Обычные экспериментальные методы включают рентгеновскую кристаллографию и ЯМР-спектроскопию, оба из которых могут давать структурную информацию с атомным разрешением. Однако эксперименты с ЯМР могут предоставить информацию, на основе которой можно оценить подмножество расстояний между парами атомов, а окончательные возможные конформации для белка определяются путем решения задачи геометрии расстояния. Интерферометрия с двойной поляризацией представляет собой количественный аналитический метод для измерения общей конформации белка и конформационных изменений из-за взаимодействий или других стимулов. Круговой дихроизм - еще один лабораторный метод определения внутреннего β-листового / α-спирального состава белков. Криоэлектронная микроскопия используется для получения структурной информации с низким разрешением об очень больших белковых комплексах, включая собранные вирусы ; вариант, известный как электронная кристаллография, в некоторых случаях также может давать информацию с высоким разрешением, особенно для двумерных кристаллов мембранных белков. Решенные структуры обычно хранятся в Protein Data Bank (PDB), свободно доступном ресурсе, из которого можно получить структурные данные о тысячах белков в форме декартовых координат для каждого атома. в белке.
Известно гораздо больше генных последовательностей, чем белковые структуры. Кроме того, набор решенных структур смещен в сторону белков, которые можно легко подвергнуть условиям, требуемым в рентгеновской кристаллографии, одном из основных методов определения структуры. В частности, глобулярные белки сравнительно легко кристаллизовать при подготовке к рентгеновской кристаллографии. Мембранные белки и большие белковые комплексы, напротив, трудно кристаллизовать и недостаточно представлены в PDB. Инициативы в области структурной геномики пытались исправить эти недостатки путем систематического определения репрезентативных структур основных классов складок. Предсказание структуры белка. Методы пытаются предоставить средства создания правдоподобной структуры для белков, структура которых не была экспериментально определена.
 Составляющие аминокислоты могут быть проанализированы для прогнозирования вторичной, третичной и четвертичной структуры белка, в данном случае гемоглобина, содержащего гем единиц
Составляющие аминокислоты могут быть проанализированы для прогнозирования вторичной, третичной и четвертичной структуры белка, в данном случае гемоглобина, содержащего гем единиц В дополнение к области структурной геномики, прогнозирование структуры белка разрабатывает эффективные математические модели белков для теоретически предсказывать молекулярные образования с помощью вычислений, а не обнаруживать структуры с помощью лабораторных наблюдений. Наиболее успешный тип предсказания структуры, известный как моделирование гомологии, основан на существовании «шаблонной» структуры с последовательностью, подобной моделируемому белку; Цель структурной геномики - обеспечить достаточное представление в решенных структурах для моделирования большинства из оставшихся. Хотя создание точных моделей остается проблемой, когда доступны только отдаленно связанные матричные структуры, было высказано предположение, что выравнивание последовательностей является узким местом в этом процессе, поскольку достаточно точные модели могут быть получены, если «идеальное» выравнивание последовательностей известен. Многие методы предсказания структуры послужили источником информации для развивающейся области белковой инженерии, в которой уже созданы новые белковые складки. Более сложной вычислительной задачей является предсказание межмолекулярных взаимодействий, например, в молекулярном стыковке и предсказании взаимодействия белок-белок.
Математические модели для моделирования динамических процессов сворачивания белка и связывание связано с молекулярной механикой, в частности, молекулярной динамикой. Методы Монте-Карло облегчают вычисления, в которых используются достижения в области параллельных и распределенных вычислений (например, проект Folding @ home, который выполняет молекулярное моделирование на графических процессорах ). Моделирование in silico обнаружило сворачивание небольших α-спиральных белковых доменов, таких как головной убор виллин и вспомогательный белок ВИЧ. Гибридные методы, сочетающие стандартную молекулярную динамику с квантовой механикой математикой, исследовали электронные состояния родопсинов.
Многие белки (у эукариот ~ 33%) содержат большие неструктурированные, но биологически функциональные сегменты и могут быть классифицированы как внутренне неупорядоченные белки. Таким образом, прогнозирование и анализ нарушения белков является важной частью характеристики структуры белка.
Большинство микроорганизмов и растения могут биосинтезировать все 20 стандартных аминокислот, тогда как животные (включая человека) должны получать некоторые аминокислоты из диеты. Аминокислоты, которые организм не может синтезировать самостоятельно, называются незаменимыми аминокислотами. Ключевые ферменты, синтезирующие определенные аминокислоты, не присутствуют у животных, такие как аспартокиназа, которая катализирует первую стадию синтеза лизина, метионина и треонин из аспартата. Если аминокислоты присутствуют в окружающей среде, микроорганизмы могут сохранять энергию, поглощая аминокислоты из окружающей среды и подавляя свои биосинтетические пути.
У животных аминокислоты получают в результате употребления продуктов, содержащих белок. Затем проглоченные белки расщепляются на аминокислоты посредством переваривания, которое обычно включает денатурацию белка посредством воздействия кислоты и гидролиз посредством ферменты, называемые протеазами. Некоторые поступившие внутрь аминокислоты используются для биосинтеза белка, в то время как другие превращаются в глюкозу через глюконеогенез или подаются в цикл лимонной кислоты. Такое использование белка в качестве топлива особенно важно в условиях голодания, поскольку оно позволяет использовать собственные белки организма для поддержания жизни, особенно те, которые содержатся в мышцах.
у животных, таких как собаки и У кошек белок поддерживает здоровье и качество кожи, способствуя росту волосяных фолликулов и кератинизации, тем самым снижая вероятность проблем с кожей, вызывающих неприятный запах. Низкокачественные белки также влияют на здоровье желудочно-кишечного тракта, увеличивая вероятность метеоризма и пахучих соединений у собак, потому что, когда белки достигают толстой кишки в непереваренном состоянии, они ферментируются с образованием сероводорода, индола и скатола. Собаки и кошки усваивают животные белки лучше, чем растительные, но продукты низкого качества животного происхождения, в том числе кожа, перья и соединительная ткань, усваиваются плохо.
Общий азот Содержание органического вещества в белках в основном формируется аминогруппами. Общий азот по Кьельдалю (TKN ) - это показатель азота, широко используемый при анализе (сточных) воды, почвы, пищевых продуктов, кормов и органических веществ в целом. Как следует из названия, применяется метод Кьельдаля. Доступны более чувствительные методы.
| На сайте Wikimedia Commons есть материалы, связанные с Proteins. |
| Найдите protein в Wiktionary, бесплатном словаре. |