
Войти
Геномика - это междисциплинарная область биологии с упором на преобразование, функцию, эволюция, картирование и редактирование геномов. Геном - это полный набор ДНК организма, включая все его гены. В отличие от генетики, которая относится к изучению отдельных генов и их роли в наследовании, геномикалена на коллективную характеристику всех генов организма, их взаимосвязей и функций на организм. Гены могут управлять производством белков с помощью ферментов и молекул-посредников. В свою очередь, белки составляют структуры тела, такие как органы и ткани, а также контролируют химические реакции и передают сигналы между клетками. Геномика также включает секвенирование и анализ геномов с использованием высокопроизводительных секвенирования ДНК и биоинформатики для сборки и анализа функций и структур целых геномов. Достижения в области геномики вызвали революцию в исследованиях, основанных на открытии, и системной биологии, чтобы облегчить понимание даже самых сложных биологических систем, таких как мозг.
Эта область также включает исследования внутригеномных (внутри генома) такие явления, как эпистаз (влияние одного гена на другого), плейотропия (один ген влияет на более чем один признак), гетерозис (сила гибрида) и другие взаимодействия между локусами и аллелями в геноме.
От греческого ΓΕΝ gen, «ген» (гамма, эпсилон, ню, эпсилон), что означает «становиться, создавать, сотворение, рождение», и последующие варианты: генеалогия, генезис, генетика, ген, геномер, генотип, род и т. Д. слово геном (от немецкого Геном, приписываемого Гансу Винклеру ) использовалось в английском еще в 1926 году, термин геномика был придуман Томом Родериком, генетик из лаборатории Джексона (Бар-Харбор, штат Мэн ), за пивом на встрече, состоявшейся в Мэриленде по картированию геном человека в 1986 году.
Последующие Розалинд Франклин подтверждает спиральную переносимость ДНК, Джеймс Д. Уотсон и Фрэнсис Крик опубликовали активность ДНК в 1953 году и После публикации Фредом Сэнгером след аминокислот инсулина в 1955 году секвенирование нуклеиновых кислот стало основной целью первых молекулярных биологов. В 1964 г. Роберт В. Холли и его коллеги опубликовали первую когда-либо определенную последовательность нуклеиновой кислоты, последовательность рибонуклеотида для транспортной РНК аланина . Расширяя эту работу, Маршалл Ниренберг и Филип Ледер раскрыли триплетную природу генетического кода и смогли определить 54 из 64 кодоны в своих экспериментах. В 1972 году Уолтер Файерс и его команда из Лаборатории молекулярной биологии Гентского университета (Гент, Бельгия ) были сначала для определения гена: гена бактериофага MS2 оболочки оболочки. Группа Файерса расширила свою работу с белками оболочки MS2, определена полная нуклеотидная последовательность бактериофага четыре MS2-РНК (чей геном кодирует всего гена в 3569 парах основ [п.н.]) и вируса обезьяны 40 в 1976 и 1978 годах соответственно.
 Фредерик Сэнджер
Фредерик Сэнджер  Уолтер Гилберт Фредерик Сэнджер и Уолтер Гилберт разделили половину лауреата Нобелевской программы по химии 1980 года за независимые методы анализа. секвенирования ДНК.
Уолтер Гилберт Фредерик Сэнджер и Уолтер Гилберт разделили половину лауреата Нобелевской программы по химии 1980 года за независимые методы анализа. секвенирования ДНК. В дополнение к его плодотворной работе по аминокислотной последовательности инсулина Фредерик Сэнгер и его коллеги сыграли ключевую роль в разработке методов секвенирования ДНК, которые позволили создать комплексные проекты секвенирования генома. В 1975 году он и Алан Коулсон опубликовал секвенирование с использованием ДНК-полимеразы с радиоактивно меченными нуклеотидами, которые он назвал техникой плюс и минус. Это включает два связанных метода, которые генерируют короткие олигонуклеотиды с определенными 3'-концами. Их можно фракционировать с помощью электрофореза на полиакриламидном геле (так называемый электрофорез в полиакриламидном геле) и визуализировать с помощью авторадиографии. Процедура могла упорядочить до 80 нуклеотидов за один раз и была большим улучшением, но все же очень очень трудоемкой. Тем не менее, в 1977 году его группа смогла секвенировать часть из 5386 нуклеотидов одноцепочечного бактериофага φX174, завершив первый полностью секвенированный геном на основе ДНК. Уточнение метода Плюс и Минус привело к обрыву цепи или методу Сенгера (см. ниже ), который лег в основу методов секвенирования ДНК, картирования генома, хранения данных и биоинформатического анализа наиболее широко использовались в следующие четверть века исследований. В том же году Уолтер Гилберт и Аллан Максам из Гарвардского университета независимо друг от друга разработали метод Максама-Гилберта (также известный как химический метод) секвенирования ДНК, предполагающего преимущественное расщепление ДНК по основаниям, менее эффективный метод. За свою новаторскую работу по секвенированию нуклеиновых кислот Гилберт и Сэнгер разделили половину Нобелевской программы 1980 года по химии с Полом Бергом (рекомбинантной ДНК ).
Появление этих технологий привело к быстрому увеличению объема и скорости выполнения проектов секвенирования генома. О первой полной последовательности генома эукариотической органеллы, задней митохондрии (16,568 п.н., около 16,6 т.п.н. [килобаз]), было сообщено в 1981 году, и о первом хлоропласте геномов последовали в 1986 году. В 1992 году была секвенирована первая эукариотическая хромосома, хромосома III пивных дрожжей Saccharomyces cerevisiae (315 кб). Первым свободноживущим организмом, который был секвенирован, был Haemophilus influenzae (1,8 МБ [мегабаза]) в 1995 году. В следующем году консорциум исследователей из лабораторий Северной Америки, Европа и Япония объявили о завершении создания первой последовательности генома эукариота, S. cerevisiae (12,1), и с тех пор секвенирование геномов продолжалось с экспоненциально растущей скоростью. По состоянию на октябрь 2011 г. полные следуют доступны для: 2719 вирусов, 1115 архей и бактерий и 36 эукариот, из которых примерно половина из них - грибы.
 Число проектов по геному увеличилось, поскольку технологические усовершенствования продолжают снижать стоимость секвенирования. (A) Экспоненциальный рост баз данных о последовательностях генома с 1995 года. (B) Стоимость в долларах США (USD) секвенирования одного миллиона оснований. (C) Стоимость в долларах США для секвенирования генома размером 3000 МБ (размером с человека) в логарифмическом масштабе.
Число проектов по геному увеличилось, поскольку технологические усовершенствования продолжают снижать стоимость секвенирования. (A) Экспоненциальный рост баз данных о последовательностях генома с 1995 года. (B) Стоимость в долларах США (USD) секвенирования одного миллиона оснований. (C) Стоимость в долларах США для секвенирования генома размером 3000 МБ (размером с человека) в логарифмическом масштабе. Большинство микроорганизмов, чьи геномы были полностью секвенированы, являются патогенами, например, Haemophilus influenzae, что привело к явному смещению их филогенетического распределения по широтой микробного разнообразия. Из других секвенированных видов большинства было выбрано, потому что они были хорошо изучены модельными организмами или обещали стать хорошими моделями. Дрожжи (Saccharomyces cerevisiae ) долгое время были важным модельным организмом для эукариотической клетки, в то время как плодовая мушка Drosophila melanogaster была очень важна инструмент (особенно в ранней домолекулярной генетике ). Червь Caenorhabditis elegans - часто используемая простая модель для многоклеточных организмов. Рыбка данио Brachydanio rerio используется для многих исследований развития на молекулярном уровне, растение Arabidopsis thaliana является модельным организмом для цветковых растений. Японский иглобрюх (Takifugu rubripes ) и пятнистый зеленый иглобрюх (Tetraodon nigroviridis ) интересны из-за их небольших и компактных геномов, которые содержат очень мало некодирующей ДНК по сравнению с большинством видов. М искусающие: собака (Canisiliaris ), бурая крыса (Rattus norvegicus ), мышь (Mus musculus ) и шимпанзе (Pan troglodytes ) являются важными модельными животными в медицинских исследованиях.
Черновой набросок генома человека был завершен Проектом генома человека в начале 2001 года, вызвав много шума. В рамках этого проекта, завершенного в 2003 году, была секвенирован весь геном одного конкретного человека, и к 2007 году эта последовательность была объявлена «завершенной» (менее одной на 20 000 оснований и собраны все хромосомы). За прошедшие с тех пор годы геномы многих других людей были секвенированы, частично под эгидой проекта Проект "1000 геномов", который объявил о секвенировании 1092 геномов в октябре 2012 года. Завершение этого проекта стало возможным разработкой значительно более эффективных технологий секвенирования и потребовало выделения значительных биоинформатических ресурсов в результате большого международного сотрудничества. Постоянный анализ геномных данных человека глубокие политические и социальные последствия для человеческого общества.
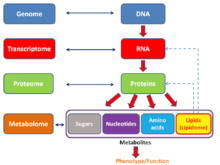 Общая схема, показывающая взаимосвязи генома, транскриптома, протеом и метаболом ( липидом ).
Общая схема, показывающая взаимосвязи генома, транскриптома, протеом и метаболом ( липидом ).Англоязычный неологизм омикс неформально относится к области биологии, оканчивающиеся на -омикс, такие как геномика, протеомика или метаболомика. Соответствующий суффикс -ome используется для обозначения объектов исследования такие поля, как геном, протеом или метаболом соответственно., используется в молекулярной биологии, относится к некоторой совокупности; аналогично омикс стал в целом в целом к изучению всеобъемлющих наборов биологических данных. В то время как рост использования этого термина привел к, что некоторые ученые (Джонатан Эйзен и другие) заявили, что это было перепродано, это отражает изменение ориентации на количественный анализ полного анализа или почти полного ассорти мента всех составляющих системы. Например, при изучении симбиозов исследователи, которые раньше ограничивались изучением одного генного продукта, теперь одновременно сравнивают набор различных типов биологических молекул.
После выбора проекта системы генома включает три компонента: секвенирование ДНК, сборку этой последовательности для создания представления исходной хромосомы, а также аннотацию и анализ этого представления.
 Обзор геномного проекта. Во-первых, необходимо выбрать геном, что включает несколько факторов, включая стоимость и актуальность. Во-вторых, генерируется и создается в данном центре секвенирования (такой как BGI или DOE JGI ). В-третьих, последовательность генома аннотируется на нескольких уровнях: ДНК, белок, генные пути или сравнительно.
Обзор геномного проекта. Во-первых, необходимо выбрать геном, что включает несколько факторов, включая стоимость и актуальность. Во-вторых, генерируется и создается в данном центре секвенирования (такой как BGI или DOE JGI ). В-третьих, последовательность генома аннотируется на нескольких уровнях: ДНК, белок, генные пути или сравнительно. Исторически секвенирование производственного процесса в центрах секвенирования, централизованных объектов (начиная с независимых организаций, таких как Объединенный институт генома, объединенный институт генома, секвенирует десятки терабазов в год на местных базовых объектах химической биологии), в исследовательских лабораториях с дорогостоящим оборудованием и необходимой технической поддержкой. Однако по мере совершенствования технологии нового поколения эффективных настольных секвенаторов с быстрым оборотом стало доступным для средней академической лаборатории. В целом подходы к секвенированию генома делятся на две большие категории: дробовик и высокопроизводительное (следующее поколение) секвенирование.
 Генетический анализатор ABI PRISM 3100. Такие капиллярные секвенаторы автоматизировали ранние крупномасштабные усилия по секвенированию генома.
Генетический анализатор ABI PRISM 3100. Такие капиллярные секвенаторы автоматизировали ранние крупномасштабные усилия по секвенированию генома. Секвенирование дробовиком - это метод секвенирования, анализа последовательностей ДНК длиной более 1000 пар оснований, вплоть до целых хромосом. Он назван по аналогии с быстро расширяющейся квазислучайной схемой стрельбы из дробовика . Секвенирование с помощью гель-электрофореза можно использовать только для некоторых коротких последовательностей (от 100 до 1000 пар оснований), более длинные последовательности ДНК должны быть разбиты на случайные небольшие сегменты, которые секвенируют для считываний. Множественные перекрывающиеся чтения цели ДНК путем выполнения нескольких раундов фрагментации и секвенирования. Затем компьютерные программы используют перекрывающиеся концы различных операций чтения, чтобы собрать их в непрерывную последовательность. Секвенирование дробовиком - это процесс случайной выборки, требующий избыточной выборки, чтобы данный нуклеотид представлен в реконструированной последовательности; среднее количество считываний, при которых происходит избыточная выборка генома, называется охват.
На протяжении большей части истории технология, лежащая в основе секвенирования дробовика, была классическим методом обрыва цепи или «методом Сэнгера ', который основан на избирательном включении завершающих цепей дидезоксинуклеотидов с помощью ДНК-полимеразы во время in vitro репликации ДНК. В последнее время на секвенировании «дробовиком» пришли методы смены высокопроизводительного секвенирования, особенно для крупномасштабных автоматизированных анализов генома. Метод Сэнгера по-прежнему широко используется, в первую очередь для небольших проектов и для получения длинных считываний непрерывных последовательностей ДНК (>500 нуклеотидов). Для методов терминации цепи требуются одноцепочечная ДНК-матрица, ДНК праймер, ДНК-полимераза, нормальные дезоксинуклеозидетрифосфаты (dNTP) и модифицированные нуклеотиды (дидезоксиNTP), останавливающие удлинение цепи ДНК. В этих обрывающих цепь нуклеотидах отсутствует группа 3'- OH, необходимая для образования фосфодиэфирной связи между двумя нуклеотидами, в результате чего ДНК-полимераза прекращает удлинение ДНК при ddNTP. DdNTP могут быть радиоактивно или флуоресцентно помечены для обнаружения в секвенаторах ДНК. Как правило, эти машины могут секвенировать до 96 образцов ДНК за одну серию (прогон) за 48 прогонов в день.
Высокопроизводительное секвенирование стимулирует развитие технологий высокопроизводительного секвенирования, которые распараллеливают процесс секвенирования, создавая тысячи или миллионы последовательностей одновременно. Высокопроизводительное секвенирование предназначено для снижения стоимости секвенирования ДНК по сравнению со стандартными методами определения терминатора с использованием красителя. При сверхвысокопроизводительном секвенировании можно выполнять до 500 000 операций секвенирования путем синтеза.
 Анализатор генома системы Illumina II. Технологии Illumina установили стандарт высокопроизводительного массового параллельного секвенирования.
Анализатор генома системы Illumina II. Технологии Illumina установили стандарт высокопроизводительного массового параллельного секвенирования. Метод секвенирования красителей Illumina основан на использовании обратимых терминаторов красителей был разработан в 1996 году в Женевском институте биомедицинских исследований Паскалем Майером. и Лоран Фаринелли. В этом методе молекулы ДНК и праймеры сначала прикрепляются к предметному стеклу и амплифицируются с помощью полимеразы, так что образуются локальные клональные колонии, используемые названные «колониями ДНК». Для определения последовательности четыре типа оснований обратимых терминаторов (RT-основания) и смывают невключенные нуклеотиды. В отличие от пиросеквенирования цепи ДНК удлиняются один нуклеотид за раз, и получение изображения может быть обработано с задержкой, что позволяет захватывать очень большие массивы колоний ДНК с помощью последовательных изображений, полученных с одной камеры. Разделение ферментативной реакции и захвата изображения обеспечивает оптимальную производительность и теоретически неограниченную емкость секвенирования; при оптимальной конфигурации максимальная производительность прибора зависит только от скорости аналого-цифрового преобразования камеры. Камера делает снимки флуоресцентно меченных нуклеотидов, затем краситель вместе с концевым блокатором 3 'химически удаляется из ДНК, обеспечивая следующий цикл.
Альтернативный подход, Ионно-полупроводниковое секвенирование основано на стандартной химии репликации ДНК. Эта технология измеряет высвобождение иона водорода каждый раз, когда вводится основание. Микролунка, содержащая матричную ДНК, заполняется одним нуклеотидом, если нуклеотид комплементарен матричной цепи, он будет включен, и ион водорода будет высвобожден. Этот выпуск запускает ионный датчик ISFET. Если гомополимер присутствует в матричной последовательности, несколько нуклеотидов будут включены в один цикл заполнения, и обнаруженный электрический сигнал будет пропорционально выше.
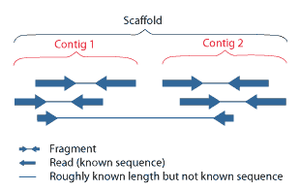 Перекрывающиеся считывания из контигов ; контиги и пробелы известной длины образуют каркасы.
Перекрывающиеся считывания из контигов ; контиги и пробелы известной длины образуют каркасы. 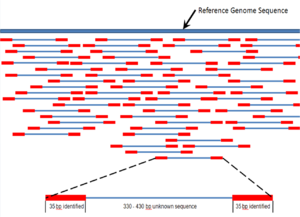 Парные считывания концов данных секвенирования следующего поколения, сопоставленные с эталонным геномом. Множественные фрагментированные считывания последовательностей должны быть собраны вместе на основе их перекрывающихся областей.
Парные считывания концов данных секвенирования следующего поколения, сопоставленные с эталонным геномом. Множественные фрагментированные считывания последовательностей должны быть собраны вместе на основе их перекрывающихся областей. Последовательность сборка относится к выравниванию и слиянию фрагментов гораздо более длинной последовательности ДНК с целью восстановления исходной последовательности. Это необходимо, поскольку современная технология секвенирования ДНК не может считывать целые геномы как непрерывную последовательность, а скорее считывает небольшие фрагменты от 20 до 1000 оснований, в зависимости от используемой технологии. Технологии секвенирования третьего поколения, такие как PacBio или Oxford Nanopore, обычно генерируют чтения секвенирования длиной>10 т.п.н. однако у них высокий уровень ошибок - примерно 15 процентов. Обычно короткие фрагменты, называемые чтениями, являются результатом секвенирования дробовиком геномной ДНК или транскриптов генов (EST ).
Сборку можно в общих чертах разделить на два подхода: сборка de novo для геномов, которые не похожи на какие-либо последовательности, секвенированные в прошлом, и сравнительная сборка, при которой в качестве эталона во времясборки используется существующая последовательность близкородственного организма. сравнительная сборка, сборка de novo, сложная в вычислительном отношении (NP-hard ), что делает ее менее подходящей для технологий NGS с коротким считыванием. В рамках парадигмы сборки de novo существуют две основные стратегии сборки: стратегии Эйлера, и стратегии консенсуса-перекрытия (OLC). Стратегии OLC в конечном итоге создать гамильтонов путь через граф перекрытия, что является NP-трудной проблемой. Стратегии эйлерова пути вычислительно более управляемым, потому что они пытаются найти эйлеров путь через деБрюйн график.
Завершенные геномы, выполняющие одну непрерывную последовательность без неоднозначностей, представляющую каждый репликон.
Сборка последовательности ДНК сама по себе не имеет большого значения без дополнительного анализа. Аннотации генома - это процесс присоединения биологической информации к последовательностям, который состоит из трех основных шагов:
Инструменты автоматического аннотации выполнить это шаги in silico, в отличие от ручного аннотирования (также известное как курирование), который включает в себя человеческий опыт и потенциальную экспериментальную проверку. В идеале эти подходы сосуществуют и дополняют друг друга в одной аннотации трубопровод (также см. ниже ).
Традиционно базовым уровнем аннотации является использование BLAST для поиска сходств, а затем аннотирование геномов на основе гомологов. Совсем недавно на платформу аннотаций добавленная дополнительная информация. Дополнительная информация позволяет ручным аннотаторам деконволютировать несоответствие между генами, которыми даны одинаковые аннотации. Некоторые базы данных используют контекстную информацию генома, оценки сходства, экспериментальные данные и интеграцию других ресурсов предоставления аннотаций генома через свой подход «Подсистемы». Другие базы данных (например, Ensembl ) полагаются как на тщательно отобранные источники данных, так и на ряд программных инструментов в своем конвейере автоматической аннотации генома. Структурная аннотация состоит из идентификации геномных элементов, в первую очередь ORF и их локализации или структуры гена. Функциональная аннотация состоит из присоединения биологической информации к геномным элементам.
Необходимость воспроизводимости и эффективного управления большим объемом данных, связанных с проектомми генома, означает, что вычислительные конвейеры имеют важное применение в геномике. 35>
Функциональная геномика - это область молекулярной биологии, заключающаяся в попытке использовать огромное количество данных, полученных в ходе геномных проектов. (например, проекты секвенирования генома ) для описания функций и взаимодействий гена (и белка ). Функциональная геномика фокусируется на динамических функциях, таких как ген транскрипция, трансляция и белок-белковые взаимодействия, в отличие от статических геномной информации, таких как последовательность ДНК или структуры. Функциональная геномика пытается ответить на вопросы о функциях ДНК на уровне генов, транскриптов РНК и белковых продуктов. Ключевой характеристикой функциональных методов является их общегеномный подход к этим вопросам, как правило, с использованием высокопроизводительных методов, а не более традиционного подхода «ген за геном».
Основная отрасль геномиков по поручениям с секвенированием геномов различных организмов, но знание полных геномов создало возможности для области функциональной геномики, в основном касается паттернов экспрессии гена в различных условиях. Наиболее важными инструментами здесь являются микроматрицы и биоинформатика.
 Пример структуры структуры белка, Среднезападным центром структурной геномики.
Пример структуры структуры белка, Среднезападным центром структурной геномики. Структурная геномика ищет для описания 3-мерной структуры каждого белка, кодируемого данных геномом. Этот основанный на геноме подход позволяет использовать высокопроизводительные методы определения структуры за счет комбинации экспериментального и модельного подходов. Принципиальное различие между структурной геномикой и традиционным структурным методом заключается в том, что структурная геномика пытается определить каждого белка, кодируемого геномом, а не сосредотачивается на одном конкретном белке. При наличии полногеномных последовательностей предсказание структуры может быть выполнено быстрее за счет комбинации экспериментального и модельного подходов, потому что наличие большого количества секвенированных геномов и ранее решенных белковых структур позволяет ученым моделировать структуру на структурех ранее решенных. гомологи. Структурная геномика включает в себя использование большого количества подходов к определению структур, включая экспериментальные методы с использованием геномных последовательностей или подходов, основанные на моделировании, основанные на последовательности или структурной гомологии с белком определенной структуры или на основе химических и физических принципов для языка., не имеющий гомологии ни с одной известной структурой. В отличие от стандартной структурной биологии, определение структуры белка с помощью структурной геномики часто (но не всегда) происходит до того, как что-либо известно о функции белка. Это ставит новые задачи в структурной биоинформатике, т. Е. Определение функции белка по его 3D структура.
Эпигеномика - это исследование полного набора эпигенетических модификаций генетического материала клетки, известных как эпигеном. Эпигенетические модификации - это обратимые модификации ДНК или гистонов, которые влияют на экспрессию генов без изменений ДНК (Russell 2010, стр. 475). Двумя наиболее характерными эпигенетическими модификациями являются метилирование ДНК и модификация гистона. Эпигенетические конструкции играют роль в экспрессии и регуляции генов и участвуют во многих клеточных процессах, таких дифференцировка / развитие и туморогенез. Изучение эпигенетики на глобальном уровне стало возможным благодаря адаптации геномных высокопроизводительных анализов.
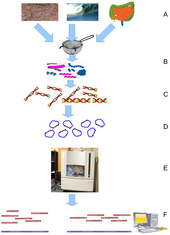 Экологическое секвенирование (ESS) - ключевой метод в метагеномике. (A) Отбор проб из среды обитания; (B) фильтрация частиц, обычно по размеру; (C) лизис и выделение ДНК; (D) клонирование и создание библиотеки; (E) секвенирование клонов; (F) сборка установить в контиги и каркасы.
Экологическое секвенирование (ESS) - ключевой метод в метагеномике. (A) Отбор проб из среды обитания; (B) фильтрация частиц, обычно по размеру; (C) лизис и выделение ДНК; (D) клонирование и создание библиотеки; (E) секвенирование клонов; (F) сборка установить в контиги и каркасы. Метагеномика - это исследование метагеномов, генетического материала, выделенного непосредственно из образцов окружающей среды. Это широкое поле может также называться экологической геномикой, экогеномикой или геномикой сообщества. В то время как традиционная микробиология и секвенирование микробного генома основываются на культивируемых клональных культурах, при раннем секвенировании генов окружающей среды клонируются специфические гены ( часто 16S рРНК ген) для получения профиля разнообразия в естественном образце. Такая работа показала, что подавляющее большинство микробного биоразнообразия было упущено с помощью методов культивирования. В недавних исследованиях использовалось «ружье» секвенирование по Сэнгеру или массовое параллельное пиросеквенирование для достижения степени объективных выборок всех генов от всех выбранных сообществ. Благодаря своей способности раскрывать ранее скрытое разнообразие микроскопической жизни, метагеномика предлагает мощную линзу для наблюдения за микробным миром, которая может революционизировать понимание всего живого мира.
Бактериофаги игрыли и продолжают играть ключевую роль в бактериальной генетике и молекулярной биологии. Исторически они использовались для определения структуры гена и регуляции гена. Также первым секвенированным геномом был бактериофаг . Однако исследования бактериофагов не приводят к революции в геномике, в которой доминирует бактериальная геномика. Лишь совсем недавно изучение геномов бактериофагов стало популярным, что исследователям удалось понять механизмы, лежащие в основе эволюции фага. Последовательности генома бактериофага могут быть получены путем прямого секвенирования бактериофагов, но также могут быть получены как часть микробных геномов. Анализ бактериальных геномов показал, что значительная часть микробной ДНК состоит из последовательностей профага и профагоподобных элементов. Подробный анализ данных последовательностей в базе данных представляет собой представление роли профагов в формировании бактериального генома: в целом, этот метод подтвердил многие известные группы бактериофагов, что делает его полезный инструмент для прогнозирования взаимосвязи профагов с бактериофагами.
В настоящее время существует 24 цианобактерий, для которых доступна полная последовательность генома. 15 из этих цианобактерий происходят из морской среды. Это шесть штаммов Prochlorococcus, семь штаммов морских Synechococcus, Trichodesmium erythraeum IMS101 и Crocosphaera watsonii. Несколько исследований могут быть использованы для вывода важных экологических и физиологических характеристик морских цианобактерий. Однако в настоящее время ведется еще много проектов по геному, которые есть еще изоляты Prochlorococcus среди морских Synechococcus, Acaryochloris и Prochloron, N 2 -фиксирующие нитчатые цианобактерии и Lyngbya majuscula, а также бактериофаги, заражающие морские цианобактерии. Таким образом, растущий объем информации о геноме можно использовать в более общем плане для решения глобальных проблем, использует сравнительный подход. Некоторыми новыми и захватывающими примерами прогресса области идентификация генов регуляторных РНК, понимание этой эволюционного происхождения фотосинтеза или оценка вклада горизонтального переноса генов в геномы, которые были проанализированы.
Геномика нашла применение во многих других областях, включая медицину, биотехнологию, антропологию и общественные науки.
Геномные технологии нового поколения позволяют клиницистам и биомедицинским исследователям значительно увеличить объем геномных данных, собираемых на больших исследуемых популяциях. В сочетании с новыми информатическими подходами, которые объединяют многие виды данных с геномными данными в исследованиях болезней, это позволяет исследователям лучше понять генетические основы реакции и болезни. Например, исследовательская программа «Все мы» направлена на сбор данных о системе от 1 миллиона участников, стать важнейшим компонентом исследовательской платформы точной медицины.
Рост Геномные знания позволили получить все более изощренные применения синтетической биологии. В 2010 году исследователи из Дж. Институт Крейга Вентера объявил о создании частично синтетического вида бактерии, Mycoplasma labratorium, происходящего из генома Mycoplasma genitalium.
Специалисты по охране природы могут использовать информацию, собранную путем геномного секвенирования, для более точной оценки генетических факторов, имеющих ключевое значение для сохранения вида, таких как генетическое разнообразие популяции или гетерозиготность особи для рецессивного наследственного генетического заболевания. Используя геномные данные для оценки эффектов эволюционных процессов и выявления закономерностей в вариациях в данной популяции, защитники природы могут сформулировать планы помощи данному виду, не оставляя неизвестных стольких переменных, как те, которые не рассматриваются стандартом генетические подходы.