
Войти
 Структура альфа-аминокислоты в ее неионизированной форме
Структура альфа-аминокислоты в ее неионизированной форме Амино кислоты - это органические соединения, содержащие амин (–NH 2) и карбоксильные (–COOH) функциональные группы вместе с боковой цепью (группа R), специфичной для каждой аминокислоты. Ключевыми элементами аминокислоты являются углерод (C), водород (H), кислород (O) и азот (N), хотя в боковых цепях некоторых аминокислот присутствуют и другие элементы. Известно около 500 встречающихся в природе аминокислот (хотя только 20 присутствуют в генетическом коде ), и их можно классифицировать по-разному. Их можно классифицировать в соответствии с положением основных структурных функциональных групп как альфа- (α-), бета- (β-), гамма- (γ-) или дельта- (δ-) аминокислот; другие категории относятся к уровню полярности, pH и типу группы боковой цепи (алифатическая, ациклическая, ароматическая, содержащая гидроксил или сера и т. д.). В форме белков аминокислотные остатки образуют второй по величине компонент (вода - самый большой) человеческих мышц и других ткани. Помимо своей роли остатков в белках, аминокислоты участвуют в ряде процессов, таких как транспорт нейротрансмиттеров и биосинтез.
В биохимии аминокислоты, которые имеют аминогруппу присоединенные к атому (альфа-) углерода рядом с карбоксильной группой, имеют особое значение. Они известны как 2-, альфа- или α-аминокислоты (в большинстве случаев общая формула H2NCHRCOOH, где R представляет собой органический заместитель, известный как «боковая цепь »); часто термин «аминокислота» используется для обозначения именно этих. Они включают 22 протеиногенных («строящих белок») аминокислот, которые объединяются в пептидные цепи («полипептиды»), чтобы сформировать строительные блоки огромного множества белки. Это все L-стереоизомеры («левосторонние » изомеры ), хотя несколько D -аминокислот («правосторонние») встречаются в бактериальных оболочках, как нейромодулятор (D-серин ), и в некоторых антибиотиках.
Двадцать протеиногенных аминокислот кодируются непосредственно триплетом кодоны в генетическом коде и известны как «стандартные» аминокислоты. Два других («нестандартный» или «неканонический») - это селеноцистеин (присутствует во многих прокариотах, а также в большинстве эукариот, но не кодируется непосредственно ДНК ) и пирролизин (обнаружен только у некоторых архей и одной бактерии ). Пирролизин и селеноцистеин кодируются через вариантные кодоны; например, селеноцистеин кодируется стоп-кодоном и элементом SECIS. N-формилметионин (который часто является исходной аминокислотой белков в бактериях, митохондрии и хлоропласты ) обычно рассматривается как форма метионина, а не как отдельная протеиногенная аминокислота. Комбинации кодон– тРНК, не встречающиеся в природе, также можно использовать для «расширения» генетического кода и образования новых белков, известных как аллопротеины, включающих не- протеиногенные аминокислоты.
Многие важные протеиногенные и непротеиногенные аминокислоты выполняют биологические функции. Например, в человеческом мозге глутамат (стандартная глутаминовая кислота ) и гамма-аминомасляная кислота («ГАМК», нестандартная гамма-аминокислота) представляют собой, соответственно, основные возбуждающие и тормозные нейротрансмиттеры. Гидроксипролин, главный компонент соединительной ткани коллаген, синтезируется из пролин. Глицин является биосинтетическим предшественником порфиринов, используемых в красных кровяных тельцах. Карнитин используется в транспорте липидов.
Девять протеиногенных аминокислот называются «незаменимыми » для человека, потому что они не могут быть произведены из других соединений путем человеческое тело и поэтому должно приниматься в пищу. Другие могут быть условно необходимыми для определенного возраста или состояния здоровья. Незаменимые аминокислоты могут также различаться между видами.
Из-за их биологического значения аминокислоты важны для питания и обычно используются в пищевых добавках, удобрениях, корма и пищевая техника. Промышленное использование включает производство лекарств, биоразлагаемых пластиков и хиральных катализаторов.
Первые несколько аминокислот были открыты в начале 19 века. В 1806 году французские химики Луи-Николя Воклен и Пьер Жан Робике выделили в спарже соединение, которое впоследствии было названо аспарагином, первым аминокислота должна быть открыта. Цистин был открыт в 1810 году, хотя его мономер, цистеин, оставался не обнаруженным до 1884 года. Глицин и лейцин были открыты в 1820 году. Последней из 20 общих аминокислот, которые были обнаружены, был треонин в 1935 году Уильямом Каммингом Роузом, который также определил незаменимые аминокислоты и установил минимальные суточные потребности всех аминокислот для оптимального роста.
Единство химической категории было признано Вюрцем в 1865 году, но он не дал ей особого названия. Первое использование термина «аминокислота» на английском языке датируется 1898 годом, тогда как немецкий термин Aminosäure использовался раньше. Было обнаружено, что белки дают аминокислоты после ферментативного переваривания или кислотного гидролиза. В 1902 году Эмиль Фишер и Франц Хофмайстер независимо друг от друга предположили, что белки образуются из многих аминокислот, посредством чего связи образуются между аминогруппой одной аминокислоты с карбоксильной группой другой, приводя к линейной структуре, которую Фишер назвал «пептидом ".
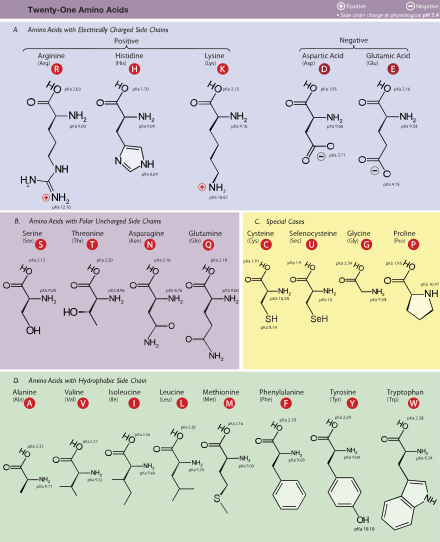 21 протеиногенная α-аминокислота, обнаруженная в эукариотах, сгруппированных по их боковым цепям 'pKa значения и заряды, переносимые при физиологическом pH (7,4)
21 протеиногенная α-аминокислота, обнаруженная в эукариотах, сгруппированных по их боковым цепям 'pKa значения и заряды, переносимые при физиологическом pH (7,4) В структуре, показанной вверху страницы, R представляет собой боковую цепь, специфичную для каждой аминокислоты. Атом углерода рядом с карбоксильной группой (которая, следовательно, имеет номер 2 в углеродной цепи, начиная с этой функциональной группы) называется α – углерод. Аминокислоты, содержащие аминогруппу, связанную непосредственно с альфа-углеродом, называются альфа-аминокислотами. К ним относятся такие аминокислоты, как пролин, которые вместе ntain вторичные амины, которые раньше часто назывались «иминокислоты».
Альфа-аминокислоты являются наиболее распространенной формой, встречающейся в природе, но только когда встречается в L -изомере. Альфа-углерод представляет собой хиральный атом углерода, за исключением глицина, который имеет два неотличимых атома водорода на альфа-углероде. Следовательно, все альфа-аминокислоты, кроме глицина, могут существовать в любом из двух энантиомеров, называемых L или D аминокислотами (относительная конфигурация)., которые являются зеркальным отображением друг друга (см. также Хиральность ). В то время как L -аминокислоты представляют собой все аминокислоты, обнаруженные в белках во время трансляции в рибосоме, D -аминокислоты обнаруживаются в некоторых белках, продуцируемых ферментом. посттрансляционные модификации после транслокации и транслокации в эндоплазматический ретикулум, как у экзотических морских организмов, таких как конусные улитки. Они также являются обильными компонентами пептидогликана клеточных стенок бактерий, а D -серин может действовать как нейромедиатор в головном мозге. D -аминокислоты используются в рацемической кристаллографии для создания центросимметричных кристаллов, которые (в зависимости от белка) могут позволить более простое и надежное определение структуры белка. Условные обозначения L и D для конфигурации аминокислот относятся не к оптической активности самой аминокислоты, а скорее к оптической активности изомера глицеральдегида из что эта аминокислота теоретически может быть синтезирована (D -глицеральдегид является правовращающим; L -глицеральдегид является левовращающим). В качестве альтернативы, обозначения (S) и (R) используются для указания абсолютной конфигурации. Почти все аминокислоты в белках являются (S) у α-углерода, причем цистеин является (R), а глицин не- хиральным. Цистеин имеет боковую цепь в том же геометрическом положении, что и другие аминокислоты, но терминология R / S перевернута, поскольку более высокий атомный номер серы по сравнению с карбоксильным кислородом дает боковой цепи более высокий приоритет по правилам Кана-Ингольда-Прелога, тогда как атомы в других боковых цепях придают им более низкий приоритет по сравнению с карбоксильной группой.
Аминокислоты обозначаются как α-, когда атом азота присоединен к атому углерода, соседнему с карбоксильной группой: в этом случае соединение содержит субструктуру N – C – CO 2. Аминокислоты с субструктурой N – C – C – CO 2 классифицируются как β-аминокислоты. γ-Аминокислоты содержат субструктуру N – C – C – C – CO 2 и т. д.
Аминокислоты обычно классифицируются по свойствам их боковую цепь на четыре группы. Боковая цепь может превращать аминокислоту в слабую кислоту или слабое основание и гидрофил, если боковая цепь полярная или гидрофоб, если он неполярный. химические структуры 22 стандартных аминокислот, наряду с их химическими свойствами, более подробно описаны в статье об этих протеиногенных аминокислотах.
Фраза «амино с разветвленной цепью кислоты "или BCAA относятся к аминокислотам, имеющим алифатические боковые цепи, которые являются линейными; это лейцин, изолейцин и валин. Пролин является единственной протеиногенной аминокислотой, боковая группа которой связана с α-аминогруппой, и, таким образом, также является единственной протеиногенной аминокислотой, содержащей вторичный амин в этом положении. Таким образом, с химической точки зрения пролин представляет собой иминокислоту, поскольку в нем отсутствует первичная аминогруппа, хотя он по-прежнему классифицируется как аминокислота в современной биохимической номенклатуре и может также называться «N-алкилированная альфа-аминокислота».
 Аминокислота в ее (1) молекулярной и (2) цвиттерионной формах
Аминокислота в ее (1) молекулярной и (2) цвиттерионной формах В водном растворе аминокислоты существуют в двух формах (как показано справа), молекулярная форма и цвиттерион находятся в равновесии друг с другом. Две формы сосуществуют в диапазоне pH от pK 1 - 2 до pK 2 + 2, который для глицина составляет pH 0–12. Соотношение концентраций двух изомеров не зависит от pH. Значение этого отношения не может быть определено экспериментально.
Поскольку все аминокислоты содержат функциональные группы аминов и карбоновых кислот, они являются амфипротонными. При pH = pK 1 (приблизительно 2,2) будет равная концентрация компонентов NH +. 3CH (R) CO. 2H и NH +. 3CH (R) CO-. 2 и при pH = pK 2 (приблизительно 10) будет равная концентрация компонентов NH +. 3CH (R) CO-. 2 и NH. 2CH (R) CO-. 2. Отсюда следует, что нейтральная молекула и цвиттер-ион являются фактически единственными частицами, присутствующими при биологическом pH.
Обычно предполагается, что концентрация цвиттер-иона намного больше, чем концентрация нейтральной молекулы на основе сравнений. с известными значениями pK аминов и карбоновых кислот.
 Состав кривых титрования двадцати протеиногенных аминокислот, сгруппированных по категории боковых цепей
Состав кривых титрования двадцати протеиногенных аминокислот, сгруппированных по категории боковых цепей При значениях pH между двумя значениями pK a цвиттерион преобладает, но сосуществует в динамическом равновесии с небольшими количествами чистых отрицательных и чистых положительных ионов. В точной средней точке между двумя значениями pK a следовое количество чистых отрицательных и следовых чистых положительных ионов точно уравновешивается, так что средний чистый заряд всех присутствующих форм равен нулю. Этот pH известен как изоэлектрическая точка pI, поэтому pI = 1/2 (pK a1 + pK a2). Для аминокислот с заряженными боковыми цепями участвует pK a боковой цепи. Таким образом, для аспартата или глутамата с отрицательными боковыми цепями pI = 1/2 (pK a1 + pK a (R)), где pK a (R) представляет собой боковую цепь pK a. Цистеин также имеет потенциально отрицательную боковую цепь с pK a (R) = 8,14, поэтому pI следует рассчитывать как для аспартата и глутамата, даже если боковая цепь не имеет значительного заряда при физиологическом pH. Для гистидина, лизина и аргинина с положительными боковыми цепями pI = 1/2 (pK a (R) + pK a2). Аминокислоты имеют нулевую подвижность при электрофорезе в их изоэлектрической точке, хотя это поведение чаще используется для пептидов и белков, чем для отдельных аминокислот. Цвиттерионы имеют минимальную растворимость в их изоэлектрической точке, и некоторые аминокислоты (в частности, с неполярными боковыми цепями) могут быть выделены путем осаждения из воды путем доведения pH до требуемой изоэлектрической точки.
 A полипептид представляет собой неразветвленную цепь аминокислот
A полипептид представляет собой неразветвленную цепь аминокислот 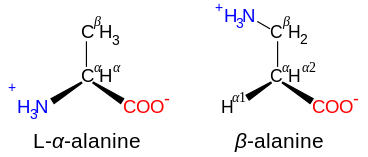 β-аланин и его изомер α-аланина
β-аланин и его изомер α-аланина 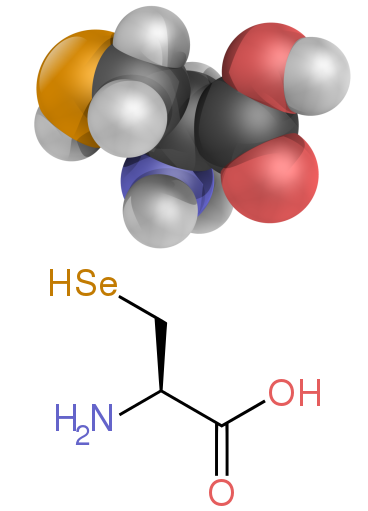 Аминокислота селеноцистеин
Аминокислота селеноцистеин Аминокислоты представляют собой структурные единицы (мономеры ), из которых состоят белки. Они соединяются вместе с образованием коротких полимерных цепей, называемых пептидами, или более длинных цепей, называемых либо полипептидами, либо белками. Эти полимеры являются линейными и неразветвленными, каждая аминокислота в цепи присоединена к двум соседним аминокислотам. Процесс создания белков, кодируемых генетическим материалом ДНК / РНК, называется трансляцией и включает в себя поэтапное добавление аминокислот к растущей белковой цепи с помощью рибозима, который называется рибосома. Порядок, в котором добавляются аминокислоты, считывается с помощью генетического кода из мРНК матрицы, которая является копией РНК одной из гены.
Двадцать две аминокислоты естественным образом включены в полипептиды и называются протеиногенными или природными аминокислотами. Из них 20 кодируются универсальным генетическим кодом . Оставшиеся 2, селеноцистеин и пирролизин, включаются в белки с помощью уникальных синтетических механизмов. Селеноцистеин включается, когда транслируемая мРНК включает элемент SECIS, который заставляет кодон UGA кодировать селеноцистеин вместо стоп-кодона . Пирролизин используется некоторыми метаногенными археями в ферментах, которые они используют для производства метана. Он кодируется кодоном UAG, который обычно является стоп-кодоном у других организмов. За этим кодоном UAG следует нижележащая последовательность PYLIS.
Помимо 22 протеиногенных аминокислот, известно множество непротеиногенных аминокислот. Они либо не обнаруживаются в белках (например, карнитин, ГАМК, левотироксин ), либо не производятся напрямую и изолированно стандартными клеточными механизмами (например, гидроксипролин и селенометионин ).
Непротеиногенные аминокислоты, которые обнаруживаются в белках, образуются с помощью посттрансляционной модификации, которая представляет собой модификацию после трансляции во время синтеза белка. Эти модификации часто важны для функции или регуляции белка. Например, карбоксилирование глутамата позволяет лучше связывать катионы кальция, а коллаген содержит гидроксипролин, образованный гидроксилированием. из пролин. Другим примером является образование гипузина в факторе инициации трансляции EIF5A посредством модификации остатка лизина. Такие модификации могут также определять локализацию белка, например, добавление длинных гидрофобных групп может вызвать связывание белка с мембраной фосфолипида.
Некоторые непротеиногенные аминокислоты не являются содержится в белках. Примеры включают 2-аминоизомасляную кислоту и нейротрансмиттер гамма-аминомасляную кислоту. Непротеиногенные аминокислоты часто встречаются в качестве промежуточных продуктов в метаболических путях для стандартных аминокислот - например, орнитин и цитруллин входят в цикл мочевины., часть катаболизма аминокислоты (см. Ниже). Редким исключением из доминирования α-аминокислот в биологии является β-аминокислота бета-аланин (3-аминопропановая кислота), которая используется в растениях и микроорганизмах при синтезе пантотеновой кислоты. (витамин B 5), компонент кофермента A.
Хотя D -изомеры редко встречаются в живых организмах, грамицидин представляет собой полипептид, состоящий из смеси D - и L -аминокислот. Другими соединениями, содержащими D -аминокислоты, являются тироцидин и валиномицин. Эти соединения разрушают стенки бактериальных клеток, особенно грамположительных бактерий. По состоянию на 2011 год из 187 миллионов проанализированных аминокислот в базе данных Swiss-Prot было обнаружено только 837 D -аминокислот.
20 аминокислот, которые содержат непосредственно кодонами универсального генетического кода, называются стандартными или каноническими аминокислотами. Модифицированная форма метионина (N-формилметионин ) часто включается вместо метионина в исходной форме аминокислоты белков в бактериях, митохондриях и хлоропластах. Остальные аминокислоты называют нестандартными или неканоническими. Большинство нестандартных аминокислот также не являются протеиногенными (т.е. не могут быть включены в белки во время трансляции), но из них протеиногенными, так как они могут трансляционно трансляционно в белки, используя информацию, не закодированную в универсальном генетическом коде.
Двумя нестандартными протеиногенными аминокислотами являются селеноцистеин (присутствует во многих неэукариотах, а также среди эукариот, но не кодируется непосредственно ДНК) и пирролизин (обнаружено только у некоторых архей и одной бактерии ). Включение этих нестандартных аминокислот происходит редко. Например, 25 человеческих белков включают селеноцистеин в своей первичной структуре, структурно охарактеризованные ферменты (селеноферменты) используют селеноцистеин в качестве каталитической составляющей в своих активных центрах. Пирролизин и селеноцистеин кодируются вариантами кодонами. Например, селеноцистеин кодируется стоп-кодоном и через SECIS.
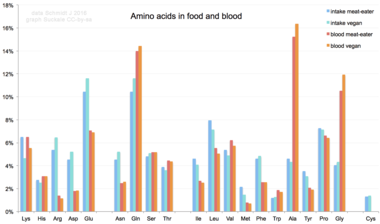 Доля аминокислот в различных диетах человека и получаемая смесь аминокислот в сыворотке крови человека. Глутамат и глутамин чаще всего присутствуют в пище - более 10%, в то время как аланин, глутамин и глицин - наиболее распространены в крови.
Доля аминокислот в различных диетах человека и получаемая смесь аминокислот в сыворотке крови человека. Глутамат и глутамин чаще всего присутствуют в пище - более 10%, в то время как аланин, глутамин и глицин - наиболее распространены в крови. При поступлении в организм из рациона используются 20 стандартных аминокислот. для синтеза белков, других биомолекул или окисляются до мочевины и диоксида углерода в качестве источника энергии. Путь окисления начинается с удаления аминогруппы трансаминазой ; аминогруппа затем вводится в цикл мочевины. Другимтом трансамидирования является кетокислота, которая входит в цикл лимонной кислоты. Глюкогенные аминокислоты также могут быть преобразованы в глюкозу посредством глюконеогенеза. Из 20 стандартных аминокислот девять (His, Ile, Leu, Lys, Met, Phe, Thr, Trp и Val ) называются незаменимыми аминокислотами, поскольку человеческое тело не может синтезировать их из соединений на уровне, необходимом для нормального роста, поэтому они должны быть получены из пищи. Кроме того, у детей цистеин, тирозин и аргинин считаются полуосновными аминокислотами, а таурин - полуосновной аминосульфоновой кислотой. Метаболические пути, которые синтезируются мономеры, еще не полностью разработаны. Необходимые также зависят от возраста и состояния здоровья человека, поэтому трудно сделать общие выводы о диетических потребностях в некоторых аминокислотах. Диетическое воздействие нестандартной аминокислоты BMAA было связано с нейродегенеративными заболеваниями человека, включая БАС.
 Диаграмма молекулярных сигнальных каскадов, которые участвуют в миофибриллах синтез мышечного белка и митохондриальный биогенез в ответ на физические упражнения и аминокислоты или их производные (в первую очередь L-лейцин и HMB ). Многие аминокислоты, полученные из пищевого белка, способствуют активации mTORC1 и увеличивают синтез белка за счет передачи сигналов через Rag GTPases.. Сокращения и обозначения: • PLD: фосфолипаза D. • PA: фосфатидная кислота. • mTOR: механистическая мишень рапамицина. • АМФ: аденозинмонофосфат. • АТФ: аденозинтрифосфат. • AMPK: АМФ-активированная протеинкиназа. • PGC - 1α: рецептор, активируемый пролифератором пероксисом, гамма-коактиватор-1α. • S6K1: киназа p70S6. • 4EBP1: фактор инициации трансляции эукариот 4E-связывающий белок 1. • eIF4E: фактор инициации трансляции эукариот 4E. • RPS6: рибосомный белок S6. • eEF2: фактор элонгации эукариот 2. • RE: упражнения с сопротивлением; ЭЭ: упражнения на выносливость. • Мио: миофибриллярные ; Мито: митохондрии. • AA: аминокислоты. • HMB: β-гидрокси β-метилмасляная кислота. • ↑ означает активацию. • Τ означает ингибирование
Диаграмма молекулярных сигнальных каскадов, которые участвуют в миофибриллах синтез мышечного белка и митохондриальный биогенез в ответ на физические упражнения и аминокислоты или их производные (в первую очередь L-лейцин и HMB ). Многие аминокислоты, полученные из пищевого белка, способствуют активации mTORC1 и увеличивают синтез белка за счет передачи сигналов через Rag GTPases.. Сокращения и обозначения: • PLD: фосфолипаза D. • PA: фосфатидная кислота. • mTOR: механистическая мишень рапамицина. • АМФ: аденозинмонофосфат. • АТФ: аденозинтрифосфат. • AMPK: АМФ-активированная протеинкиназа. • PGC - 1α: рецептор, активируемый пролифератором пероксисом, гамма-коактиватор-1α. • S6K1: киназа p70S6. • 4EBP1: фактор инициации трансляции эукариот 4E-связывающий белок 1. • eIF4E: фактор инициации трансляции эукариот 4E. • RPS6: рибосомный белок S6. • eEF2: фактор элонгации эукариот 2. • RE: упражнения с сопротивлением; ЭЭ: упражнения на выносливость. • Мио: миофибриллярные ; Мито: митохондрии. • AA: аминокислоты. • HMB: β-гидрокси β-метилмасляная кислота. • ↑ означает активацию. • Τ означает ингибирование  Тренировка с отягощениями стимулирует синтез мышечного протеина (MPS) в течение периода до 48 часов после тренировки (показано более светлым пунктиром). Прием богатой белком пищей в любой момент в течение этого периода увеличитанное упражнениями увеличение синтез мышечного белка (показано сплошными линиями).
Тренировка с отягощениями стимулирует синтез мышечного протеина (MPS) в течение периода до 48 часов после тренировки (показано более светлым пунктиром). Прием богатой белком пищей в любой момент в течение этого периода увеличитанное упражнениями увеличение синтез мышечного белка (показано сплошными линиями). Биосинтетические пути для катехоламинов и следы аминов в головном мозге человека  L-фенилаланин L-тирозин L-ДОФА эпинефрин фенэтиламин р-тирамин допамин Норэпинефрин N-Метилфенэтиламин N-Метилтирамин п-Октопамин Синефрин 3-Метокситирамин AADC AADC AADC первичный. путь PNMT PNMT PNMT PNMT AAAH AAAH мозг. CYP2D6 второстепенный. путь COMT DBH DBH L-фенилаланин L-тирозин L-ДОФА эпинефрин фенэтиламин р-тирамин допамин Норэпинефрин N-Метилфенэтиламин N-Метилтирамин п-Октопамин Синефрин 3-Метокситирамин AADC AADC AADC первичный. путь PNMT PNMT PNMT PNMT AAAH AAAH мозг. CYP2D6 второстепенный. путь COMT DBH DBH |
У людей небелковые аминокислоты также играют роль, как промежуточные продукты метаболизма, такие как биосинтез нейротрансмиттера гамма-аминомасляной кислоты (ГАМК). Многие аминокислоты используются для использования других молекул, например:
Некоторые нестандартные аминокислоты используются в качестве защиты от травоядных у растений. Например, канаванин является аналогом аргинина, который содержится во многих бобовых, и в особенно больших количествах в Canavalia gladiata (меч фасоль). Эта аминокислота защищает растения от хищников, как насекомые, вызывающие болезни у людей, если некоторые виды бобовых употребляются в пищу без обработки. Небелковая аминокислота мимозин обнаружена в других лучших бобовых, в частности в Leucaena leucocephala. Это соединение является аналогом тирозина и может отравлять животных, которые пасутся на этих растениях.
Аминокислоты используются в различных отраслях промышленности, но в основном они используются в качестве добавок к корму для животных. Это необходимо, поскольку многие из основных компонентов этих кормов, такие как соевые бобы, имеют низкие уровни, либо не имеют некоторых незаменимых аминокислот : лизина, метионина, треонина и триптофан играет важную роль роль в производстве этих кормов. В этой промышленности аминокислоты также используются для хелатирования катионов металлов с целью улучшения абсорбции минералов из добавок, которые могут потребоваться для улучшения здоровья или продуктивности этих животных.
пищевая промышленность. также является основным потребителем аминокислот, в частности, глутаминовой кислоты, которая используется в качестве усилителя вкуса, и аспартама (аспартилфенилаланин 1-метиловый эфир) в качестве низкокалорийного искусственного подсластителя. Используется в индустрии питания человека для облегчения симптомов дефицита минералов, путем улучшения всасывания минералов и негативных побочных эффектов от добавок неорганических минералов.
Хелатирующая способность аминокислоты использовалась в удобрениях для сельского хозяйства, чтобы облегчить доставку минералов к растениям, чтобы исправить недостаток минералов, например хлороз железа. Эти удобрения также используются для предотвращения возникновения недостатков и улучшения состояния растений. Оставшееся производство аминокислот используется в синтезе лекарств и косметики.
. Аналогичным образом, некоторые производные аналоги используются в фармацевтической промышленности. К ним защищ 5-HTP (5-гидрокситриптофан), использованный для экспериментального лечения депрессии, L-DOPA (L-дигидроксифенилаланин) для лечения болезни Паркинсона и эфлорнитин препарат, который ингибирует орнитиндекарбоксилазу и используется для лечения сонной болезни.
С 2001 года в белок было добавлено 40 неприродных аминокислот путем создания уникального кодона ( перекодирования) и пары трансфер-РНК: аминоацил-тРНК-синтетаза для кодирования различных физико-химических и биологических свойств, чтобы использовать его в качестве инструмента для исследования структуры и функции белка или для создания новых или улучшенных белков.
Нулломеры - это кодоны, которые теоретически кодируют аминокислоты, в природе существуют избирательное предубеждение против использования этого кодона в пользу другого, например, бактерии предпочитают использовать CGA вместо AGA для кодирования аргинина. Это некоторые устанавливают, которые не появляются в геноме. Эта характеристика может быть использована для создания новых селективных лекарств для борьбы с раком и для предотвращения перекрестного заражения образцов ДНК при расследованиях на месте преступления.
Аминокислоты - это важно как дешевое сырье. Эти соединения используются в синтезе хирального пула в энантиомерно чистых строительных блоков.
Аминокислоты были исследованы в качестве предшественников хиральных катализаторов, таких как для асимметричных факторов гидрирования, хотя коммерческого применения не существует.
Аминокислоты считаются компонентами биоразлагаемых полимеров, которые имеют применение в окружающей среде упаковка и в медицине доставка лекарств и изготовление протезных имплантатов. Интересным примером таких материалов является полиаспартат, водорастворимый биоразлагаемый полимер, который может найти применение в одноразовых подгузниках и в сельском хозяйстве. Благодаря своей растворимости и способности хелатировать ионы, полиаспартат также используется в качестве биоразлагаемого агента против образования накипи и ингибитора коррозии. Кроме того, ароматическая аминокислота тирозин рассматривается как возможная замена фенолов, таких как бисфенол A, при производстве поликарбонатов.
 Синтез аминокислот по Стрекеру
Синтез аминокислот по Стрекеру Коммерческое производство аминокислот обычно зависит от мутантных бактерий, производящих избыточное количество отдельных аминокислот, используя глюкозу в качестве источника углерода. Некоторые аминокислоты производятся путем ферментативного превращения синтетических промежуточных продуктов. 2-Аминотиазолин-4-карбоновая кислота является промежуточным продуктом, например, в одном промышленном синтезе L-цистеина. Аспарагиновая кислота образуется добавлением аммиака к фумарату с использованием лиазы.
В растениях азот сначала ассимилируется в органические соединения в форме глутамата, образованного из альфа-кетоглутарата и аммиака в митохондрии. Что касается других аминокислот, растения используют трансаминазы для перемещения аминогруппы от глутамата к другой альфа-кетокислоте. Например, аспартатаминотрансфераза превращает глутамат и оксалоацетат в альфа-кетоглутарат и аспартат. Другие организмы также используют трансаминазы для синтеза аминокислот.
Нестандартные аминокислоты обычно образуются в результате модификаций стандартных аминокислот. Например, гомоцистеин образуется посредством пути транссульфирования или путем деметилирования метионина через промежуточный метаболит S-аденозилметионин, тогда как гидроксипролин образуется в результате посттрансляционной модификации пролина.
Микроорганизмов, а растения синтезируют много необычных аминокислот. Например, некоторые микробы производят 2-аминоизомасляную кислоту и лантионин, который является производным аланина с сульфидной мостиковой связью. Обе эти аминокислоты обнаружены в пептидных лантибиотиках, таких как аламетицин. Однако в растениях 1-аминоциклопропан-1-карбоновая кислота представляет собой небольшую дизамещенную циклическую аминокислоту, которая является ключевым промежуточным продуктом в производстве растительного гормона этилен.
Аминокислоты подвергаются реакциям, ожидаемым от составляющих функциональных групп.
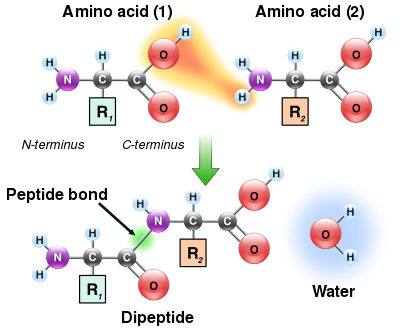 Конденсация двух аминокислот с образованием дипептида через пептидную связь
Конденсация двух аминокислот с образованием дипептида через пептидную связь Поскольку как аминовая, так и карбоксильная группы аминокислот могут реагировать с образованием амидных связей, одна молекула аминокислоты может реагировать с другой и соединяться посредством амидной связи. Эта полимеризация аминокислот создает белки. Эта реакция конденсации дает вновь образованную пептидную связь и молекулу воды. В клетках эта реакция не происходит напрямую; вместо этого аминокислота сначала активируется присоединением к молекуле транспортной РНК через связь сложного эфира. Эта аминоацил-тРНК продуцируется в АТФ -зависимой реакции, осуществляемой аминоацил-тРНК-синтетазой. Эта аминоацил-тРНК затем является субстратом для рибосомы , которая катализирует атаку аминогруппы удлиненной белковой цепи на сложноэфирную связь. В результате этого механизма все белки, производимые рибосомами, синтезируются, начиная с их N-конца и продвигаясь к их C-концу.
Однако не все пептидные связи образуются таким образом. В некоторых случаях пептиды синтезируются специфическими ферментами. Например, трипептид глутатион является важной частью защиты клеток от окислительного стресса. Этот пептид синтезируется в два этапа из свободных аминокислот. На первой стадии гамма-глутамилцистеинсинтетаза конденсирует цистеин и глутаминовую кислоту через пептидную связь, образованную между карбоксилом боковой цепи глутамата (гамма-углерод эту боковую цепь) и аминогруппу цистеина. Затем этот дипептид конденсируется с глицином с помощью глутатионсинтетазы с образованием глутатиона.
В химии пептиды синтезируются с помощью различных реакций. Один из наиболее часто используемых в твердофазном пептидном синтезе использует ароматические оксимные производные аминокислот в качестве активированных единиц. Их добавляют последовательно к растущей пептидной цепи, которая прикреплена к твердой смоляной подложке. Библиотеки пептидов используются при открытии лекарств посредством высокопроизводительного скрининга.
Комбинация функциональных групп позволяет аминокислотам быть эффективными полидентатными лигандами для хелатов металл-аминокислота. Множественные боковые цепи аминокислот также могут вступать в химические реакции.
 Катаболизм протеиногенных аминокислот. Аминокислоты могут быть классифицированы по свойствам их основных продуктов как одно из следующих:. * Глюкогенные, с продуктами, имеющими способность образовывать глюкозу посредством глюконеогенеза. * Кетогенный, при этом продукты не обладают способностью образовывать глюкозу. Эти продукты все еще можно использовать для кетогенеза или синтеза липидов.. * Аминокислоты катаболизируются как в глюкогенные, так и в кетогенные продукты.
Катаболизм протеиногенных аминокислот. Аминокислоты могут быть классифицированы по свойствам их основных продуктов как одно из следующих:. * Глюкогенные, с продуктами, имеющими способность образовывать глюкозу посредством глюконеогенеза. * Кетогенный, при этом продукты не обладают способностью образовывать глюкозу. Эти продукты все еще можно использовать для кетогенеза или синтеза липидов.. * Аминокислоты катаболизируются как в глюкогенные, так и в кетогенные продукты. Аминокислоты должны сначала выйти из органов и клеток в кровоток, переносок через аминокислоты, поскольку группы аминов и карбоновых кислот обычно являются ионизированным. Распад аминокислоты, происходящий в печени и почках, часто включает >за перемещение аминогруппы в альфа-кетоглутарат с образованием глутамата. В этом процессе участвуют трансаминазы, которые используются при аминировании во время синтеза. У многих аминогруппа затем удаляется через цикл мочевины и выводится в форму мочевины. Однако разложение аминокислот может привести к образованию мочевой кислоты или аммиака. Например, сериндегидратаза превращает серин в пируват и аммиак. После удаления одной или нескольких аминогрупп остаток молекулы иногда можно использовать для синтеза новых аминокислот или для получения энергии, введя гликолиз или цикл лимонной кислоты, как показано на изображении справа.
Аминокислоты замещают собой бидентатные лиганды, образующие аминокислоты комплексы переходных металлов.
Приблизительно. 20 канонических аминокислот можно классифицировать по их свойствам. Важными факторами являются заряд, гидрофильность или гидрофобность, размер и функциональные группы. Эти свойства воздействия на изменение белка и белок-белковые взаимодействия. Водорастворимые белки обычно имеют гидрофобные остатки (Leu, Ile, Val, Phe и Trp ) погребен в середине белка, подвернувшиеся воздействию водного растворителя. (Обратите внимание, что в биохимии остаток относится к конкретному мономеру в полимерной цепи полисахарида, белка или нуклеиновая кислота.) интегральные мембранные белки, как правило, имеют внешние кольца открытых гидрофобных аминокислот, которые закрепляют их в липидном бислое. Некоторые белки периферической мембраны имеют на своей поверхности участок гидрофобных аминокислот, который фиксируется на мембране. Аналогичным образом белки, которые должны связываться с положительно заряженными молекулами, имеют поверхности, богатые отрицательно заряженными аминокислотами, такими как глутамат и аспартат, в то время как белки, связывающиеся с отрицательно заряженными молекулами, имеют поверхности, богатые положительно заряженными аминокислотами. заряженные цепи, такие как лизин и аргинин. Существуют различные шкалы гидрофобности аминокислотных остатков.
Некоторые аминокислоты обладают особыми свойствами, такими как цистеин, которые могут образовывать ковалентные дисульфидные связи к другим остаткам цистеина, пролин, который образует цикл с основной цепью полипептида, и глицин, который более гибкий, чем другие аминокислоты.
Многие белки претерпевают ряд посттрансляционных модификаций, в результате чего дополнительные химические группы присоединяются к боковым цепям аминокислот. Некоторые модификации могут производить гидрофобные липопротеины или гидрофильные гликопротеины. Этот тип модификации позволяет обратимое нацеливание белка на мембрану. Например, добавление и удаление жирной кислоты пальмитиновой кислоты с остатками цистеина в некоторых сигнальных белках приводит к тому, что белки прикрепляются, а затем отделяются от клеточных мембран.
| Аминокислота | Буквенный код | Боковая цепь | Гидропатия. индекс | Молярная абсорбция | Молекулярная масса | Содержание в белках (%) | Стандартное генетическое кодирование, нотация IUPAC | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 1 | Класс | Полярность | Заряд, при pH 7,4 | Длина волны, λ макс. (нм) | Коэффициент, ε (мМ · см) | |||||
| Аланин | Ala | A | Алифатический | Неполярный | Нейтральный | 1,8 | 89,094 | 8,76 | GCN | ||
| Аргинин | Arg | R | Базовый | Базовый полярный | Положительный | -4,5 | 174,203 | 5,78 | MGR, CGY (кодоны также могут быть выражены: CGN, AGR) | ||
| аспарагин | Asn | N | Амид | Полярный | Нейтральный | −3,5 | 132.119 | 3,93 | AAY | ||
| Аспарагиновая кислота | Asp | D | Кислота | Кислотная полярная | Отрицательная | −3,5 | 133,104 | 5,49 | GAY | ||
| Цистеин | Cys | C | Серный | Неполярный | Нейтральный | 2,5 | 250 | 0,3 | 121,154 | 1,38 | UGY |
| Глютамин | Gln | Q | Амид | Полярный | Нейтральный | -3,5 | 146,146 | 3,9 | CAR | ||
| Глутаминовая кислота | Glu | E | Кислота | Кислая полярная | Отрицательная | −3,5 | 147,131 | 6,32 | GAR | ||
| Глицин | Гли | G | Алифатический | Неполярный | Нейтральный | -0,4 | 75,067 | 7,03 | GGN | ||
| Гистидин | His | H | Основной ароматический | Основной полярный | Положительный, 10%. Нейтральный, 90% | -3,2 | 211 | 5,9 | 155,156 | 2,26 | CAY |
| Изолейцин | Иле | I | Алифатический | Неполярный | Нейтральный | 4,5 | 131,175 | 5,49 | AUH | ||
| лейцин | Leu | L | алифатический | неполярный | нейтральный | 3,8 | 131,175 | 9,68 | YUR, CUY (кодирующие кодоны также могут быть выражены: CUN, UUR) | ||
| лизин | Lys | K | Basic | Базовый полярный | Положительный | -3,9 | 146,189 | 5,19 | AAR | ||
| Метионин | Met | M | Серная | Неполярная | Нейтральная | 1,9 | 149,208 | 2,32 | AUG | ||
| Фенилаланин | Phe | F | Ароматический | Неполярный | Нейтральный | 2,8 | 257, 206, 188 | 0,2, 9,3, 60,0 | 165,192 | 3,87 | UUY |
| Proline | Pro | P | Циклический | Неполярный | Нейтральный | - 1,6 | 115,132 | 5,02 | CCN | ||
| Серин | Сер | S | Гидроксильный | Полярный | Нейтральный | −0,8 | 105,093 | 7,14 | UCN, AGY | ||
| Треонин | Thr | T | Гидроксильный | Полярный | Нейтральный | -0,7 | 119,119 | 5,53 | ACN | ||
| Триптофан | Trp | W | Ароматический | Неполярный | Нейтральный | -0,9 | 280, 219 | 5,6, 47,0 | 204,228 | 1,25 | UGG |
| Тирозин | Тир | Y | Ароматический | Полярный | Нейтральный | -1,3 | 274, 222, 193 | 1,4, 8,0, 48,0 | 181,191 | 2,91 | UAY |
| валин | Val | V | алифатический | неполярный | Нейтральный | 4,2 | 117,148 | 6,73 | GUN | ||
Две дополнительные аминокислоты у некоторых видов кодируются кодонами , которые обычно интерпретируются как стоп-кодоны :
| 21-я и 22-я аминокислоты | 3-буквенные | 1-буквенные | Молекулярная масса |
|---|---|---|---|
| Селеноцистеин | Sec | U | 168.064 |
| пирролизин | Pyl | O | 255.313 |
В дополнение к конкретным кодам аминокислот, заполнители используются в случаях, когда химический или кристаллографический анализ AP Эптид или белок не могут окончательно определить идентичность остатка. Они также используются для суммирования мотивов консервативной белковой последовательности. Использование одинарных букв для обозначения наборов похожих остатков аналогично использованию кодов аббревиатуры для вырожденных оснований.
| Неоднозначные аминокислоты | 3-буквенные | 1-буквенные | Включены аминокислоты | Включены кодоны |
|---|---|---|---|---|
| Любое / неизвестно | Xaa | X | Все | NNN |
| Аспарагин или аспарагиновая кислота | Asx | B | D, N | RAY |
| Глютамин или глутаминовая кислота | Glx | Z | E, Q | SAR |
| лейцин или изолейцин | Xle | J | I, L | YTR, ATH, CTY (кодирующие кодоны также могут выражаться: CTN, ATH, TTR; MTY, YTR, ATA; MTY, HTA, YTG) |
| Гидрофобный | Φ | V, I, L, F, W, Y, M | NTN, TAY, TGG | |
| Ароматический | Ω | F, W, Y, H | YWY, TTY, TGG (кодоны также могут быть выражены: TWY, CAY, TGG) | |
| Алифатический (неароматический) | Ψ | V, I, L, M | VTN, TTR (кодоны также могут выражаться: NTR, VTY) | |
| Small | π | P, G, A, S | BCN, RGY, GGR | |
| Гидрофильный | ζ | S, T, H, N, Q, E, D, K, R | VAN, WCN, CGN, AGY (кодирующие кодоны также могут быть выражены: VAN, WCN, MGY, CGP) | |
| Положительно заряженные | + | K, R, H | ARR, CRY, CGR | |
| Отрицательно заряженный | − | D, E | GAN |
Unk иногда используется вместо Xaa, но менее стандартен.
Кроме того, многие нестандартные аминокислоты имеют определенный код. Например, некоторые пептидные препараты, такие как бортезомиб и MG132, искусственно синтезированы и сохраняют свои защитные группы, которые имеют определенные коды. Бортезомиб - это Pyz –Phe – boroLeu, а MG132 - это Z –Leu – Leu – Leu – al. Чтобы помочь в анализе структуры белка, доступны фотореактивные аналоги аминокислот. К ним относятся (pLeu ) и (pMet ).
Общее содержание азота в органическом веществе в основном формируется аминогруппами в белках. Общий азот по Кьельдалю ( TKN ) - это показатель азота, широко используемый при анализе (сточных) воды, почвы, пищевых продуктов, кормов и органических веществ в целом. Как следует из названия, метод Кьельдаля является Доступны более чувствительные методы.