Войти
| Эукариот. Временной диапазон: Орозириан - Настоящее время 1850–0 млн. Лет Фаза Протерозой Архей Хад'н | |
|---|---|
 | |
| Эукариоты и некоторые примеры их разнообразия - по часовой стрелке сверху слева: Красная пчела-каменщик, Boletus edulis, шимпанзе, Isotricha Кишечник, Ranunculus asiaticus и Volvox carteri | |
| Научная классификация | |
| Домен: | Eukaryota. (Чаттон, 1925) Уиттакер Маргулис, 1978 |
| Супергруппы и царства | |
Эукариотические организмы, которые нельзя уничтожить объединенные в царстве Plantae, Animalia или Fungi, иногда объединяются в царство Protista. | |
Eukaryotes () - организмы имеют чьи клетки ядро , заключенное в ядерную оболочку. Эукариоты принадлежат к домену Eukaryota или Eukarya ; их название происходит от греческого εὖ (eu, «колодец» или «хороший»)и κάρυον (карион, «орех» или «ядро»). Домен Eukaryota составляет одну из сфер жизни в трехдоменной системе ; два других домена - это бактерии и археи (вместе известные как прокариоты ). Эукариоты представляют крошечное меньшинство среди живых организмов; однако из-за их, как правило, намного большего размера, их совокупная всемирная биомасса оценивается примерно равной биомассе прокариот. Эукариоты возникли примерно 1,6–2,1 миллиарда летназад, в протерозойском эоне.
Эукариотические клетки обычно содержат связанные с мембраной органеллы, такие как митохондрии и аппарат Гольджи, а также хлоропласты. в растениях и водорослях ; эти органеллы уникальны для эукариот, хотя примитивные органеллы можно найти у прокариот. Помимо того, что эукариоты являются одноклеточными, они также могут быть многоклеточными и множеством типовклеток, образующих различные типы ткани ; для сравнения, прокариоты обычно одноклеточные. Животные, растения и грибы - наиболее известные эукариоты; другие эукариоты иногда называют простейшими.
Эукариоты могут воспроизводить как бесполым через митоз, так и половым путем посредством мейоза и гамет слияния. В митозе одна клетка делится, чтобы произвести две генетически идентичные клетки. В мейозе зарепликацией ДНК следуют два цикла деления клеток с образованием четырех гаплоидных дочерних клеток. Они как половые клетки (гаметы). Каждая гамета имеет только один набор хромосом, каждая из которых представляет собой уникальное сочетание комбинации родительских хромосом в результате генетической рекомбинации время мейоза.
 Константин Мерещковский использует симбиотическое происхождение клеток с ядрами.
Константин Мерещковский использует симбиотическое происхождение клеток с ядрами. Концепция эукариота была приписанафранцузскому биологу Эдуарду Шаттону (1883–1947). Термины «прокариот» и «эукариот» были более определенно введены канадским микробиологом Роджером Стэнье и голландско-американским микробиологом С. Б. ван Ниль в 1962 году. В своей работе «Titres et Travaux Scientifiques» 1937 года использует два термина, назвав бактерии прокариотами и организмы с ядрами в своих клетках эукариотами. Однако он представил об этом только в одном абзаце, и эта идея фактически игнорироваласьдо тех, пока заявление Чаттона не было открыто снова Станье и ван Нилом.
В 1905 и 1910 годах русский биолог Константин Мерещковский (1855) –1921) утверждал, что пластиды были редуцированы цианобактериями в симбиозе с не- фотосинтетическим (гетеротрофным ) хозяин, который сам был сформирован симбиозом между амебоподобным хозяином и бактериоподобной клеткой, которая сформировала ядро. Таким образом, растения унаследовали фотосинтез отцианобактерий.
В 1967 году Линн Маргулис представила микробиологические доказательства эндосимбиоза как источник хлоропластов и митохондрий в эукариотических клетках в своей статье, О происхождении митозирующих клеток. В 1970-х годах Карл Вёзе исследовал микробную филогенетику, изучая вариации в 16S рибосомной РНК. Это помогло раскрыть происхождение эукариот и симбиогенез двух важных эукариот органелл, митохондрий и хлоропластов. В 1977 году Вёзе и Джордж Фокс представили «третью форму жизни», которую они назвали архебактериями; в 1990 году Везе, Отто Кандлер и Марк Л. Уилис переименовали это в археи.
В 1979 году Г.В. Гулд и Г.Дж. Дринг предположили, что ядро эукариотической клетки произошло от способности грамположительных бактерий образовывать эндоспоры. В 1987 г. И более поздних работах Томасалье-Смит вместо этого, чтобы мембраны ядраи использовали эндоплазматический ретикулум сначала образовывались путем складывания плазматической мембраны прокариота. В 1990-х годах несколько других биологов предложили эндосимбиотическое происхождение ядра, эффективно возродив теорию Мерешковского.
 Воспроизвести медиа Видео цитологии, особенности клетки
Воспроизвести медиа Видео цитологии, особенности клетки Эукариотические клетки обычно намного больше, чем таковые из прокариот, объемом примерно в 10 000 раз больше, чемпрокариотическая клетка. Они имеют множество внутренних мембранных связующих структур, называемых органеллами, и цитоскелетом, состоящим из микротрубочек, микрофиламентов и промежуточные волокна, которые играют роль в определении и клетки формы. Эукариотическая ДНК разделена на несколько линейных пучков, называемых хромосомами, которые разделены веретеном микротрубочек во время деления ядра.
 Эндомембранная система и ее компоненты
Эндомембранная система и ее компоненты Клетки эукариотов включают в себя множество мембраносвязанных структур, вместе именуемых эндомембранной системы. Простые компартменты, называемые пузырьками и вакуолями, могут образовываться, отделяясь от других мембран. Многие клетки поглощают пищу и другие материалы в процессе эндоцитоза, когда внешняя мембрана инвагинирует, а отщипывается, образуя везикулу. Вероятно, что другиемембранно-связанные органелл в конечном итоге происходит из таких везикул. В качестве альтернативы некоторые продукты, продуцируемые клеткой, могут покидать везикулы посредством экзоцитоза.
Ядро окружено двойной мембраной (обычно называемой ядерной мембраной или ядерной оболочкой) с порами, которые позволяют материалу входить и выходить. Различные трубчатые и пластинчатые продолжения ядерной мембраны эндоплазматический ретикулум, который участвует в транспорте исозревании белка. Он включает грубый эндоплазматический ретикулум, к которым прикреплены рибосомы для белков, которые попадают во внутреннее пространство или просвет. Впервые они обычно проникают в пузырьки, которые отпочковываются от гладкой эндоплазматической сети. У больших эукариот эти несущие белок везикулы высвобождаются и модифицируются в стопках уплощенных везикул (цистерн ), аппарат Гольджи.
Везикулы могут быть специализированы для различных целей. Например,лизосомы содержат пищеварительные ферменты, которые расщепляют большинство биомолекул в цитоплазме. Пероксисомы используются для расщепления пероксида, который в остальном токсичен. Многие простейшие имеют сократительные вакуоли, которые собирают иесняют избыточную воду, и экструзомы, которые выталкивают материал, используемые для отражения хищников или поимки производства. У высших растений большую часть объема занимаетцентральная вакуоль, которая в основном содержит воду и в первую очередь поддерживает ее осмотическое давление.
 Упрощенная структура митохондрии
Упрощенная структура митохондрии Митохондрии Митохондрии обнаружены у всех эукариот, кроме одного. Митохондрии энергией эукариотическую клетку, сахар в АТФ. Они две окружающие мембраны, имеющие каждую из которых представляют собой двухслойный фосфолипид ; внутренний из которых сложен в инвагинации,называемые кристами, где происходит аэробное дыхание.
Внешняя митохондриальная мембрана является свободно проницаемой и позволяет почти всему входить в межмембранное пространство, в то время как внутренняя митохондриальная мембрана полупроницаема, поэтому пропускает только некоторые необходимые вещи в матрицу митохондрий.
Митохондрии содержат собственную ДНК, которая имеет близкое структурное сходство с бактериальной ДНК и кодирует генырРНК и тРНК, продуцирующие РНК, которые по структуре ближе к бактериальной РНК, чем к эукариотической РНК. В настоящее время принято, что они произошли от эндосимбиотических прокариот, вероятно, протеобактерий.
некоторых эукариот, таких как метамонад, таких как Giardia и Трихомонады и амебозойное Пеломикса, по-видимому, лишены митохондрий, но все они содержат органеллы, происходящие из митохондрий, такие как гидрогеносомы и митосомы и, таким образом, вторично потеряли свои митохондрии. Они получают энергию за счет ферментативного воздействия на питательные вещества, поступающие из окружающей среды. Метамонад Monocercomonoides также приобрел посредством латерального переноса гена цитозольную систему мобилизации серы, которая обеспечивает кластеры железа и серы, необходимые для синтеза белка. Нормальный митохондриальный путь кластера железо-сера утраченвторично.
Растения и различные группы водорослей также имеют пластиды. Пластиды также имеют собственную ДНК и получены из эндосимбионтов, в данном цианобактерий. Обычно имеют форму хлоропластов, которые, как и цианобактерии, содержат хлорофилл и производят органические соединения (такие как глюкоза ) посредством фотосинтеза. Остальные занимаются хранением еды. Хотя пластиды, вероятно, имели однопроисхождение, не все пластидсодержащие группы связаны. Вместо этого некоторые эукариоты получили их в результате других эндосимбиоза или проглатывания. Захват и секвестрация фотосинтетических клеток и хлоропластов происходит у многих типов современных эукариотических организмов и известна как клептопластика.
Эндосимбиотическое происхождение также было предложено для ядра и эукариотических жгутиков.
 294 разрез жгутика>Chlamydomonas reinhardt ii
294 разрез жгутика>Chlamydomonas reinhardt ii Многие эукариоты имеют длинные тонкие подвижные цитоплазматические выступы, называемые жгутиками, или аналогичные структуры, называемые ресничками. Жгутики и реснички иногда называют ундулиподиями, и они по-разному участвуют в движении, питании и ощущениях. Они состоят в основном из тубулина. Они полностью отличаются от жгутиков прокариот. Они поддерживаются пучком микротрубочек, получают из центриоли, характерно в виде девяти дублетов,окружающих два синглета. Жгутики также могут иметь волоски, или мастигонемы, и чешуйки, соединяющие мембраны и внутренние стержни. Их внутренняя часть является непрерывной с цитоплазмой клетки.
Микрофиламентные структуры, состоящие из актина и связывающих актин белков, например, α- актинин, фимбрин, филамин также присутствует в субмембранозных корковых слоях и пучках. Моторные белки микротрубочек, например, динеин или кинезин и актин, например, миозины, обеспечение динамического характера сети.
Центриоли часто присутствуют даже в клетках и группах, не имеющих жгутиков, но хвойные и цветковые растения их не имеют. Обычно они встречаются группой, дающими начало различным корням микротрубочек. Они образуют первичный компонент структуры цитоскелета и часто собираются в ходе нескольких клеточных делений, при этом один из них остается от родителя, а другой происходит отродителя. Центриоли производят веретено во время деления ядра.
Подчеркивается значение структур цитоскелета в определении формы клеток, а также то, что они являются важными компонентами миграционных факторов, таких как хемотаксис и хемокинез. Некоторые протисты имеют другие органеллы, поддерживаемые микротрубочками. К ним относятся радиолярии и Heliozoa, которые производят аксоподии, используемые для флотации или поимки добычи, игаптофиты, которые имеют своеобразную жгутикоподобная органелла, называемая гаптонема.
Клетки растений и водорослей, грибов и большинство хромальвеолатов имеют клеточную стенку, слой за пределами клеточная мембрана, обеспечивающая клетке структурную поддержку, защиту и механизм фильтрации. Клеточная стенка также предотвращает чрезмерное расширение при попадании воды в клетку.
Основными полисахаридами,составляющими первичную клеточную стенку наземных растений, являются целлюлоза, гемицеллюлоза и пектин. Микрофибриллы целлюлозы связаны гемицеллюлозными связями с образованием целлюлозно-гемицеллюлозной сети, которая встроена в пектиновую матрицу. Наиболее распространенной гемицеллюлозой в первичной клеточной стенке является ксилоглюкан.
Существует много различных типов эукариотическихклеток, хотя животные и растения являются наиболее знакомыми эукариотами и, таким образом, отличная отправная точка для понимания структуры эукариот. Однако грибы и многие протисты имеют некоторые существенные различия.
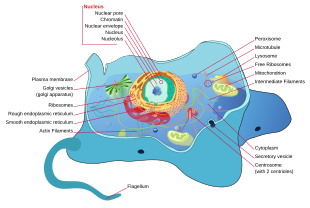 Структура типичной животной клетки
Структура типичной животной клетки 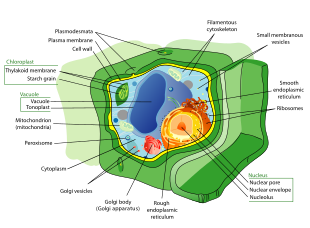 Структура типичной растительной клетки
Структура типичной растительной клетки Все животные являются эукариотами. Клетки животных отличаются от клеток других эукариот, в первую очередь растений, поскольку в них отсутствуют клеточные стенки и хлоропласты, а также имеются более мелкие вакуоли. Из-за отсутствия клеточной стенки клетки животных могут принимать различные формы. фагоцитарная клетка может даже поглощать другие структуры.
Растительные клетки сильно отличаются от клеток других эукариотических организмов. Их отличительными особенностями являются:
 Грибные клетки гиф: 1 - стенка гифы, 2 - перегородка, 3 - митохондрия, 4 -вакуоль, 5 - кристалл эргостерола, 6 - рибосома, 7 - ядро, 8 - эндоплазматический ретикулум, 9 - липидное тело, 10 - плазматическая мембрана, 11 - spitzenkörper, 12 - Аппарат Гольджи
Грибные клетки гиф: 1 - стенка гифы, 2 - перегородка, 3 - митохондрия, 4 -вакуоль, 5 - кристалл эргостерола, 6 - рибосома, 7 - ядро, 8 - эндоплазматический ретикулум, 9 - липидное тело, 10 - плазматическая мембрана, 11 - spitzenkörper, 12 - Аппарат Гольджи Клетки грибов наиболее похожи на клетки животных, за следующими исключениями:
Некоторые группы эукариот имеют уникальные органеллы, такие как цианеллы (необычные хлоропласты) глаукофитов,гаптонемы гаптофитов или эжектосомы криптомонад. Другие структуры, такие как псевдоподии, встречаются в различных группах эукариот в различных формах, таких как лобозные амебозойные или ретикулозные фораминиферы.
 Эта диаграмма иллюстрирует двойную цену секса. Если бы каждый особь принес одно и то же количество потомков (два), (а) половая популяция оставалась того же размера в каждом поколении, а (б) асексуальная популяцияудваивалась в каждом поколении.
Эта диаграмма иллюстрирует двойную цену секса. Если бы каждый особь принес одно и то же количество потомков (два), (а) половая популяция оставалась того же размера в каждом поколении, а (б) асексуальная популяцияудваивалась в каждом поколении. Деление клеток обычно происходит бесполым путем посредством митоза, процесса, который позволяет каждому дочернему ядру получить по одной копии каждой хромосомы. У большинства эукариот также есть жизненный цикл, который включает половое размножение, чередование между гаплоидной фазой, когда только одна копия каждой хромосомы присутствует в каждой клетке и диплоидная фаза, в которой две копиикаждой хромосомы присутствуют в каждой клетке. Диплоидная фаза образуется в результате слияния двух гаплоидных гамет с образованием зиготы, которая может делиться митозом или подвергаться хромосомной редукции в результате мейоза. Этот паттерн сильно различается. У животных нет многоклеточной гаплоидной фазы, но каждое поколение растений может состоять из гаплоидной и диплоидной многоклеточных фаз.
Эукариоты имеют меньшее отношение площади поверхности к объему, чем прокариоты,и, следовательно, имеют более низкую скорость метаболизма и более длительное время генерации.
Эволюция полового размножения может быть изначальной и фундаментальная характеристика эукариот. Основываясь на филогенетическом анализе, Дакс и Роджер предположили, что факультативный пол присутствовал у общего предка всех эукариот. Основной набор генов, которые функционируют в мейозе, присутствует как в Trichomonas vaginalis, так и в Giardia Кишечник, двухорганизмах, которые ранее считались бесполыми. Поскольку эти два вида являются потомками линий, которые рано разошлись с эволюционным деревом эукариот, был сделан вывод, что основные мейотические гены и, следовательно, пол, вероятно, присутствовали у общего предка всех эукариот. Было показано, что виды эукариот, которые когда-то считались бесполыми, например паразитические простейшие из рода Leishmania, имеют половой цикл. Кроме того, теперь данные показывают, что амебы, ранеесчитавшиеся бесполыми, в древности являются половыми и что большинство современных бесполых групп, вероятно, возникли недавно и независимо.
 Филогенетическое и симбиогенетическое древо живых организмов, показывает происхождение эукариот и прокариот
Филогенетическое и симбиогенетическое древо живых организмов, показывает происхождение эукариот и прокариот  Одна гипотеза эукариотических взаимоотношений - группа Opisthokonta включает как животных (Metazoa), так и грибы, растения (Plantae) помещены в Archaeplastida.
Одна гипотеза эукариотических взаимоотношений - группа Opisthokonta включает как животных (Metazoa), так и грибы, растения (Plantae) помещены в Archaeplastida. Круговая диаграмма описанных видовэукариот (кроме Excavata) вместе с деревом, показывающим возможные отношения между группами
Круговая диаграмма описанных видовэукариот (кроме Excavata) вместе с деревом, показывающим возможные отношения между группами В древности две линии происхождения животных и растения были признаны. Им был присвоен таксономический ранг из Царства от Линнея. Хотя он включил грибы с растениями с некоторыми оговорками, позже было обнаружено, что они совершенно разные и требуют отдельного царства, состав которого не был полностью ясен до 1980-х годов. Когда сталоизвестно, различные одноклеточные эукариоты изначально были помещены вместе с растениями или животными. В 1818 году немецкий биолог Георг А. Гольдфусс придумал слово простейшие для обозначения таких организмов, как инфузории, и эта группа была расширена до тех пор, пока не охватила все отдельные -клеточные эукариоты, получившие собственное царство Протиста, Эрнст Геккель в 1866 году. Таким образом, эукариоты состояли из четырех царств:
Под протистами понимались «примитивные формы» и, следовательно, эволюционная ступень, объединены своей примитивной одноклеточной природой. Распутывание глубоких расщеплений в древе жизни на самом деле началось только с секвенирования ДНК, что привело к системе доменов, а не царств в качестве ранга высшего уровня. выдвинутый Карлом Вёзе,объединяющим все царства эукариот под эукариотической областью. В то же время активизировались работы над деревом протистов, которые активно продолжаются и сегодня. Было предложено несколько альтернативных классификаций, хотя единого мнения в этой области нет.
Эукариоты - это клады, которые обычно считаются сестрой Heimdallarchaeota в группе Асгарда в архей. В одной из предложенных систем базовыми группами являются Opimoda, Diphoda, Discoba и Loukozoa. Корень эукариот обычно считается находящимся рядом или даже в Дискобе.
A Классификация, разработанная в 2005 году для Международного общества протистологов и отражающая консенсус того времени, разделила эукариот на шесть якобы монофилетических «супергрупп». Однако в том же году (2005 г.) были высказаны сомнения относительно того, были ли некоторые из этих супергрупп монофилетическими, особенно Chromalveolata, и в обзоре 2006 г. былоотмечено отсутствие доказательств для несколько из предполагаемых шести супергрупп. В пересмотренной классификации 2012 года признаются пять супергрупп.
| Archaeplastida (или Primoplantae) | Наземные растения, зеленые водоросли, красные водоросли и глаукофиты |
| SAR супергруппа | Stramenopiles (бурые водоросли, диатомеи и т. Д.), Alveolata и Rhizaria (Foraminifera, Radiolaria и различныедругие амебоидные простейшие) |
| Excavata | Различные жгутиковые простейшие |
| Amoebozoa | Most lobose амебоиды и слизистые плесени |
| Opisthokonta | Животные, грибы, хоанофлагелляты и т. д. |
Существуют также более мелкие группы эукариот положение которых не определено или, кажется, выходит за рамки основных групп, в частности, Haptophyta, Cryptophyta, Centrohelida, Telonemia, Picozoa, Apusomonadida, Ancyromonadida, Breviatea и род Collodictyon. В целом, кажется, что, хотя прогресс был достигнут, все еще существуют очень значительные неопределенности в истории эволюции и классификации эукариот. Как сказали Роджер и Симпсон в 2009 году, «при нынешних темпах изменений в нашем понимании эукариотического древа жизни мы должны действовать с осторожностью».
В статье, опубликованной в журнале Nature Microbiology вапреле 2016 года авторы «еще раз подтвердили, что жизнь, которую мы видим вокруг себя - растения, животные, люди и другие так называемые эукариоты - составляют крошечный процент мирового биоразнообразия». Они классифицировали эукариот «на основе наследования их информационных систем в отличие от липидов или других клеточных структур». Джиллиан Ф. Бэнфилд из Калифорнийского университета в Беркли и другие ученые использовали суперкомпьютер для создания диаграммы нового древа жизни наоснове ДНК 3000 видов, включая 2072 известных вида и 1011 новых микробных организмов., чью ДНК они собрали из разных сред. Поскольку способность секвенировать ДНК стала проще, Бэнфилд и команда смогли выполнить метагеномное секвенирование - «секвенировать целые сообщества организмов одновременно и выбирать отдельные группы только на основе их генов».
Деревья рРНК, построенные в течение 1980-х и 1990-х годов, оставили большинство эукариот в неразрешеннойгруппе "кроны" (технически не являющейся истинной кроной ), которую обычно разделяли по форме митохондриальные кристы; см. коронные эукариоты. Несколько групп, в которых отсутствуют митохондрии, разветвлялись отдельно, и поэтому их отсутствие считалось примитивным; но сейчас это считается артефактом притяжения длинных ветвей, и известно, что они потеряли их вторично.
По состоянию на 2011 год широко распространено мнение, что ризарии принадлежатстраменопилам. и альвеолаты в кладе, получившей название супергруппы SAR, так что ризарии не являются одной из основных групп эукариот; также, что Amoebozoa и Opisthokonta являются монофилетическими и образуют кладу, часто называемую униконтами. Помимо этого, похоже, нет консенсуса.
Было подсчитано, что может существовать 75 различных ветвей эукариот. Большинство этих линий - протисты.
Известные размеры генома эукариот варьируются от 8,2 мегабайт ( МБ) в Babesia bovis до 112 000–220 050 мегабайт у динофлагеллат, показывая, что геном предкового эукариота претерпел значительные изменения во время своего существования. эволюция. Считается, что последним общим предком всех эукариот был фаготрофный протист с ядром, по крайней мере, одной центриолью и ресничкой, факультативно аэробными митохондриями, полом (мейоз и сингамия ), спящая киста с клеточной стенкой из хитина и / или целлюлозы и пероксисом. Позднее эндосимбиоз привел к распространению пластид в некоторых линиях.
Глобальное дерево эукариот на основе консенсуса филогенетических данных (в частности, филогеномики), редких геномных подписей и морфологических характеристик представлено в Adl et al. 2012 г. и Бурки 2014/2016 гг., Когда Cryptophyta и picozoa появились в пределах Archaeplastida. Аналогичное включение Glaucophyta, Cryptista (атакже, что необычно, Haptista ) также было сделано.
| Eukaryotes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В некоторых анализах группа Hacrobia (Haptophyta + Cryptophyta ) помещается рядом с Archaeplastida, но у других он гнездится внутри Archaeplastida. Однако несколько недавних исследований пришли к выводу, что Haptophyta и Cryptophyta не образуют монофилетическую группу. Первая может быть сестринской группой с группой SAR, вторая - с Archaeplastida ( растения в широком смысле).
Разделение эукариот на две основные клады, биконты (Archaeplastida + SAR + Excavata ) и unikonts (Amoebozoa + Opisthokonta ), происходящее от предкового двублагеллярного организма и предкового однофлагеллярного организма, соответственно, были предложены ранее. В исследовании 2012 года было получено несколько аналогичное разделение, хотя было отмечено, что термины «униконты» и «биконты» неиспользовались в первоначальном смысле.
Сильно сходящийся и конгруэнтный набор деревьев появляется у Derelle et al. (2015), Ren et al. (2016), Ян и др. (2017) и Cavalier-Smith (2015), включая дополнительную информацию, что привело к более консервативному и консолидированному дереву. Он сочетается с некоторыми результатами Кавальер-Смита для базальной Opimoda. Основными остающимися противоречиями являются корень и точное расположение Rhodophyta и биконтов Rhizaria, Haptista,Cryptista, Picozoa и Telonemia, многие из которых могут быть гибридами эндосимбиотических эукариот-эукариот. Archaeplastida приобрела хлоропласты, вероятно, в результате эндосимбиоза прокариотического предка, связанного с ныне существующими cyanobacterium, Gloeomargarita lithophora.
| Eukaryotes |
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
Томас Кавальер-Смит 2010, 2013, 2014, 2017 and 2018 places the eukaryotic tree's root between Excavata (with ventral feeding grooveподдерживается микротрубочковым корнем) и без бороздок Euglenozoa, и монофилетический Chromista, коррелировал с одним эндосимбиотическим событием поимки красных водорослей. He et al. специально поддерживает укоренение эукариотического дерева между монофилетическим Discoba (Discicristata + Jakobida ) и Amorphea - Diaphoretickes клады.
| Eukaryotes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Трехдоменное дерево и Гипотеза эоцитов
Трехдоменное дерево и Гипотеза эоцитов  Филогенетическое дерево, показывающее возможную связь между эукариотами и другими формами жизни; эукариоты окрашены в красный цвет, археи зеленый и бактерии синий
Филогенетическое дерево, показывающее возможную связь между эукариотами и другими формами жизни; эукариоты окрашены в красный цвет, археи зеленый и бактерии синий  дерево эоцитов.
дерево эоцитов. Происхождение эукариотической клетки является важной вехой в эволюции жизни, поскольку эукариоты включают всех сложные клетки и почти все многоклеточные организмы. Для поиска первых эукариотов и их ближайших родственников был использован ряд подходов. последний общий предок эукариот (LECA) является гипотетическим последним общим предком всех эукариот, которые когда-либо жили, и были m Скорее всего, биологическая популяция.
эукариот имеет набор характерных особенностей, которые отличают их от других сфер жизни, включая эндомембранную систему и уникальные биохимические пути, такие как стеран синтез. Набор белков, называемых эукариотическими сигнатурными белками (ESP), был предложен для идентификации эукариотических родственников в 2002 году: к тому времени они не имели гомологии с белками, известными в других сферах жизни, но, по-видимому, они универсальны для эукариот. Они включают ude proteins that make up the cytoskeleton, the complex transcription machinery, membrane-sorting systems, the nuclear pore, as well as some enzymes in the biochemical pathways.
The timing of this series of events is hard to determine; Knoll (2006) suggests they developed approximately 1.6–2.1 billion years ago. Some acritarchs are known from at least 1.65 billion years ago, and the possible alga Grypania has been found as far back as 2.1 billionмного лет назад. Геосифон -подобное ископаемое гриб Дискагма было обнаружено в палеопочвах возрастом 2,2 миллиарда лет.
Организованные живые структуры были обнаружены в черные сланцы палеопротерозойской франсвильской формации B в Габоне, возраст которых составляет 2,1 миллиарда лет. В то время могла возникнуть эукариотическая жизнь. Окаменелости, которые явно относятся к современным группам, начинают появляться примерно на 1,2 миллиарда лет назад, в форме красных водлей, хотя недавние исследования предполагают существование окаменелых нитчатых водорослей в бассейне Виндхья, датируемых, вероятно, 1.6. до 1,7 миллиарда лет назад.
Биомаркеры предполагают, что по крайней мере стволовые эукариоты возникли еще раньше. Присутствие стеранов в австралийских сланцах указывает на то, что эукариоты присутствовали в этих породах возрастом 2,7 миллиарда лет, хотя предполагалось, что они могли происходить из-зазагрязнения образцов..
Когда бы они ни возникли, эукариоты могли стать экологически доминирующими гораздо позже; массивный рост цинкового состава морских отложений 800 миллионов лет назад был приписан росту значительных популяций эукариот, которые преимущественно потребляют и включают цинк по сравнению с прокариотами.
В апреле 2019 биологи сообщили, что очень большой медусавирус или его родственник, возможно, был ответственен, по крайней мере частично, заэволюционное появление сложных эукариотических клеток из более простых прокариотических клеток. клетки.
ядерная ДНК и генетический аппарат эукариот больше похож на Archaea, чем Бактерии, что приводит к спорное предположение, что эукариоты должны быть сгруппированы с архим в кладе Неомура. В остальном, например, по составу мембран, эукариоты похожи на Бактерии. Было предложено три основных объяснения этого:
 Схема происхождения жизни с появлением эукариот рано, а непроизошедших от прокариот, как было предложено Ричардом Эгелем в 2012 году. Эта точка зрения подразумевает, что УЦА была относительно и сложной.
Схема происхождения жизни с появлением эукариот рано, а непроизошедших от прокариот, как было предложено Ричардом Эгелем в 2012 году. Эта точка зрения подразумевает, что УЦА была относительно и сложной. Альтернативные предложения включают:
Если предположить, что никакая другая группа не участвует, существует три преступник для бактерий, архей и эукариот, в каждом из них монофилетичен.Они обозначены от 1 до 3 в таблице ниже. Гипотеза эоцитов является модификацией гипотез 2, согласно которой археи являются парафилетическими. (Таблица и название гипотез основаны на Хариш и Курланд, 2017.)
| 1 - Две империи | 2 - Три домена | 3 - Гупта | 4 - Эоцит | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
В последние годы большинства исследователей отдавали предпочтение либо три домена (3D), либо гипотез эезацитов. Анализ рРНК поддерживает сценарий эоцитов, очевидно, с корнем эукариотов в Excavata. Кладограмма, подтверждающая гипотезу эоцитов, позиционирующая эукариот внутри архей, основанная на филогеномном анализе архей Асгарда, выглядит следующим образом:
| Proteoarchaeota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
В этой сценарии группа таксон группы TACK, которая включает Crenarchaeota (ранее называвшаяся эоцитами ), Thaumarchaeota и другие. Сообщается, что эта группа содержит набор сигнатурных белков эукариот и производитвезикулы.
В 2017 году был отмечен значительный протест против этой сценария, утверждая, что эукариоты не возникли внутри архей. Cunha et al. подтверждают анализы, подтверждающие три области (3D) или гипотезу Вёза (2 в таблице выше) и отвергающие гипотезу эоцитов (4 выше). Хариш и Курланд нашел сильную поддержку двух предыдущих империй (2D) или гипотезы Майра (1 в таблице выше), сценария на аналитических кодирующих последовательностях белковых доменов. Они отвергли гипотезу эоцитов какназвание гипотезы. Возможная интерпретация их анализа состоит в том, что универсальный общий предок (UCA) нынешнего древа жизни был сложным эволюционным механизмом, который пережил узкое место, а не более простым механизмом, возникшим в начале истории жизни. С другой стороны, исследователи, которые придумали Асгард, подтвердили свою гипотезу дополнительными образцами Асгарда.
Подробная информация о взаимоотношениях представителей архей Асгарда и эукариот сообщалось, что Candidatus Prometheoarchaeum syntrophicum, тип культивируемых архей Асгарда, может быть возможной связью между простыми прокариотическими и сложными эукариотическими микроорганизмами около двух миллиардов лет назад <. 375>
Происхождение эндомембранной системы и митохондрий также неясно. фаготрофная гипотеза предполагает, что мембраны эукариотического типа, лишенные клеточной стенки, возникли первыми, с развитиемэндоцитоза, тогда как митохондрии были приобретены путем проглатывания в виде эндосимбионтов. синтрофическая гипотеза предполагает, что протоэукариотеза полагается на прото-митохондрии в качестве пищи, и поэтому в конечном итоге выросло, чтобы окружить его. Здесь мембраны возникли после поглощения митохондрии, отчасти благодаря митохондриальным генам (водородная гипотеза - одна из версий).
В исследовании с использованием геномов для построения супердеревьев, Pisani et al. (2007) предполагают, что присутствуют доказательства того, что никогда не существовало эукариот без митохондрий, эукариоты произошли от синтрофии между археями, связанными с Thermoplasmatales и α- proteobacterium, вероятно, симбиоз, вызванный серой или водородом. Митохондрия и ее геном - это остатки α-протеобактериального эндосимбионта. Большинство генов симбионта перенесено в ядро. Они составляют большую часть метаболических иэнергетических путей эукариотической системы, сохраняется у архей.
Были предложены различные гипотезы относительно того, как появились эукариотические клетки. Эти гипотезы можно разделить на два разных класса - автогенные модели и химерные модели.
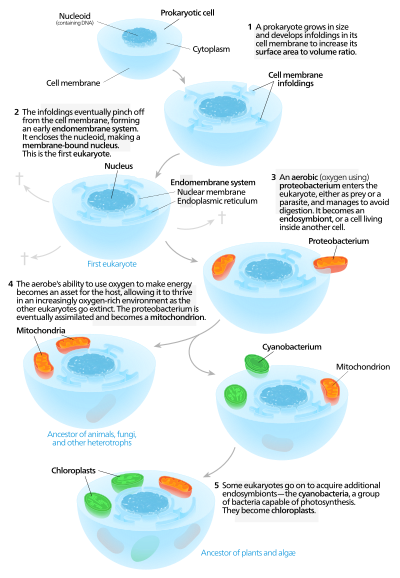 Аутогенные модели происхождения эукариот.
Аутогенные модели происхождения эукариот. Аутогенные модели предполагают, что протоэукариотическая, содержащая ядро , существовала первая, а позже приобрела митохондрии. Согласно этой модели, большой прокариот развил инвагинации в плазматической мембране, чтобы получить достаточную площадь для своего обслуживания цитоплазматический объем. По мере, как инвагинации дифференцировались по функциям, некоторые из них становились отдельными компартментами, давая начало эндомембранной системе, включая эндоплазматический ретикулум, аппарат Гольджи, ядерную мембрану.,и одинарные мембранные структуры, такие как лизосомы.
Митохондрии, предположительно происходит из эндосимбиоза аэробной протеобактерии, и решительно, что все эукариотические линии, которые не приобрели митохондрии, вымерли, и это заявление подверглось критике за отсутствие опровержения. Хлоропласты возникли в результате другого эндосимбиотического события с участием цианобактерий. Все известные эукариоты имеют митохондрии, но не все имеютхлоропласты, теория серийного эндосимбиоза предполагает, что митохондрии были первыми.
Химерные модели утверждают, что изначально существовали две прокариотические клетки - архея и бактерия. Ближайшими из ныне живущих родственников этих животных являются Asgardarchaeota и (дальние родственники) alphaproteobacteria. Эти клетки подверглись процессуального слияния либо путем физического слияния, либо посредством эндосимбиоза, что привело к образованию эукариотической клетки. В рамках этих химических моделей некоторые исследования также утверждают, что митохондрии произошли от бактериального предка, в то время как другие роли эндосимбиотических процессов в происхождении митохондрий.
Гипотеза наизнанку, разработанная двоюродными братьями Дэвид и Базз Баум, предполагает, что произошло влияние свободноживущих митохондриоподобных бактерий и архея в эукариотическуюклетку. постепенно в течение длительного периода времени вместо фагоцитоза одним глотком. В этой сценарии архей будет ловить аэробные бактерии с помощью клеточных выступов, а затем их жизнь, чтобы получать от них энергию, а не переваривать их. На ранних стадиях бактерии все еще частично находятся в прямом контакте с окружающей средой, и археону не нужно обеспечивать их необходимыми питательными веществами. В конечном итоге архея полностью поглотит бактерии, создавая в процессевнутренние мембранные структуры и мембрану ядра.
Предполагается, что группа архей, называемая галофилами, прошла через аналогичную систему, где они приобрели целую тысячу генов от бактерии, намного больше, чем посредством обычного горизонтального переноса генов, который часто происходит в микробном мире, но эти два микроба снова разделились, прежде чем они слились в одном эукариот - как клетка.
Основываясь на процессе мутуалистического симбиоза,гипотезы можно разделить на - серийную эндосимбиотическую гипотезу или теорию (SET), водородная гипотеза (в основном процесс симбиоза, при котором перенос водорода происходит между различными видами) и гипотрофии . Эти гипотезы обсуждаются отдельно в следующих разделах.
Расширенная версия гипотезы «наизнанку» предполагает, что эукариотическая клетка была создана в результате взаимодействия между двумя прокариотическими организмами ипоследний общий предок эукариот получил свой геном от всей популяции или сообщества микробов, участвующих в отношениях сотрудничества для процветания и сотрудничества. выживания в своей среде. Геномы различных типов микробов дополняют друг друга, и случайный горизонтальный перенос генов между ними будет в степени в их пользу. Это накопление полезных генов привело к возникновению генома эукариотической клетки, который содержал все гены, необходимые для независимости.
Согласно теории серийных эндосимбиотиков (отстаивает Линн Маргулис ), объединение подвижной анаэробной бактерии (например, Spirochaeta) и термоацидофильной crenarchaeon (например, Thermoplasma, которая является сульфидогенной по своей природе) дало начало сегодняшнему дню. эукариоты. Этот союз создал подвижный организм, способный жить в уже употреблении кислых и сернистых водых. Известно, что кислород токсичен для организмов, укоторых отсутствует необходимый метаболический механизм. Таким образом, архея предоставила бактерии очень благоприятную восстановленную среду (сера и сульфат были восстановлены до сульфида). В микроаэрофильных условиях кислород был восстановлен до воды, тем самым создавая платформу взаимной выгоды. Бактерия, с другой стороны, внесла необходимые продукты ферментации и акцепторы электронов вместе со своей особенностью подвижности для архей, тем самым получившим подвижность для организма.
Из консорциума бактериальной и архейной ДНК произошел ядерный геном эукариотических клеток. Спирохеты дали начало подвижным свойствам эукариотических клеток. Эндосимбиотические объединения предков аль-протеобактерий и цианобактерий вызывают к возникновению митохондрий и пластид соответственно. Например, известно, что тиодендрон возник в результате эктосимбиотического процесса, основанного на аналогичной синтрофии серы, существующей между двумя типами бактерий - Desulphobacter и Spirochaeta.
Однако такая ассоциации, основанной на подвижном симбиозе, практически никогда не наблюдалось. Также нет свидетельств того, что археи и спирохеты приспосабливаются к интенсивным кислотным средам.
В гипотезе водорода симбиотическая связь анаэробного и автотрофного метаногенная архея (хозяин) с альфа-протеобактерией (симбионт) дала начало эукариотам. Хозяиниспользовал водород (H2) и диоксид углерода (CO. 2) для производства метана, в то время как симбионт, способный к аэробному дыханию, вытеснил H 2 и CO. 2 как побочные продукты анаэробного процесса ферментации. Метаногенная среда хозяина работала как поглотитель H 2, что приводит к усиленной бактериальной ферментации.
Перенос эндосимбиотического гена (EGT) действовал как катализатор для хозяина, чтобы усвоить углеводный метаболизм симбионтов и сделать гетеротрофным по своей природе. Впечатляющая способность к образованию метана была потеря хозяна. Таким образом, происхождениееротрофной органеллы (симбионта идентично происхождение эукариотической линии . В этой гипотезе присутствие H 2 представляет собой селективную силу, которая выковала эукариот из прокариот.
Гипотеза синтрофии была выделена разница от водородная гипотеза и предполагает наличиедвух симбиотических событий. Согласно этой теории, происхождение эукариотических клеток было основано на метаболическом симбиозе (синтрофии) между метаногенными археонами и дельта-протеобактериями. Первоначально этому синтрофному симбиозу способствовал перенос H 2 между разными видами в анаэробной среде. На более ранних стадиях альфа-протеобактерия стала членом этой интеграции, превратилась в митохондрию. Перенос гена от дельта-протеобактерии к археоне привел к тому, чтометаногенная архея превратилась в ядро. Археи составляли генетический аппарат, дельта-протеобактерии вносили вклад в цитоплазматические особенности.
Эта теория включает две селективные силы во время ядра эволюции
Питтс и Гальбанон запускает сложный сценарий из 6+ серийных эндосимбиотических событий архей и бактерий, вызывающих поздней стадии эукариогенеза, возможных комбинаций, как вторичный эндосимбионт. Результаты были отвергнуты как артефакт.
![]() Эта статья включает материалы общества достояния из документа NCBI : «Science Primer».
Эта статья включает материалы общества достояния из документа NCBI : «Science Primer».
| Wikispecies содержат информацию, относящуюся к Eukaryota |