
Войти
 Фермент глюкозидаза превращает сахар мальтозу на два глюкозы сахара. Активный центр остатки красного цвета, мальтозный субстрат черным цветом и НАД кофактора желтым. (PDB : 1OBB )
Фермент глюкозидаза превращает сахар мальтозу на два глюкозы сахара. Активный центр остатки красного цвета, мальтозный субстрат черным цветом и НАД кофактора желтым. (PDB : 1OBB )Ферменты- это белки, которые включают как биологические катализаторы (биокатализаторы). Катализаторы ускоряют химические реакции. Молекулы, которые могут действовать ферменты, называются субстратами, а фермент преобразует субстраты в различные молекулы, как продукты. Почти все метаболические процессы в клетке нуждаются в ферментном катализе, чтобы происходить с достаточно высокой скоростью для поддержания жизни. Метаболические пути зависит от ферментов, которыеопределяют этапы аминокислот исследуют и необычных «псевдокаталитиче ских» свойствах.
Изве стно, что ферменты катализируют более 5000 биохимических воздействий., называемые рибозимами. Специфичность ферментов проистекает из их уникальных трехмерных структур. Р.
Как и все катализаторы, ферменты увеличивают скорость реакции за счет снижения их активации энергии. Некоторые ферменты могут превратить субстрата в продукт во много миллионовраз. Ярким примером является оротидин-5'-фосфатдекарбоксилаза, который позволяет за миллисекунды протекать реакцию, которая в силе заняла бы миллионы лет. В химическом отношении фермы любого катализатора, они не расходуются в химических реакциях и не изменяют равновесие реакции. Ферменты отличаются от других катализаторов гораздо более специфичными. На активность фермента могут влиять другие молекулы: ингибиторы - это молекулы, которые снижают активность фермента,а активаторы - это молекулы, повышающие активность. Многие терапевтические лекарства и яды являются ингибиторами ферментов. Активность фермента заметно снижается за пределами оптимальной температуры и pH, и многие ферменты (навсегда) денатурируются при воздействии чрезмерного тепла, теряя свою устойчивость и каталитические свойства.
Некоторые ферменты используются в коммерческих целях, например, при синтезе антибиотиков. Некоторые товары для дома используются ферменты для ускорения химических факторов: ферменты в биологических стиральных порошках расщепляют пятна от белка, крахмала или жира на одежде, а ферменты в размягчителе мяса расщепляют белки на более мелкие молекулы, благодаря чему легче жевать.
 Эдуард Бюхнер
Эдуард Бюхнер К концу 17 - началу 18 веков переваривание мяса секрецией желудка и превращением крахмала в сахара растительными экстрактами и слюной были известны, но механизмы, с помощью которых это происходило, не были идентифицированы.
Французский химик Ансельм Пайен был первым, кто открыл фермент диастазу в 1833 году. Несколько десятилетий спустя, изучая ферментацию сахара на спирт с помощью дрожжей, Луи Пастер пришел к выводу, что это брожение было вызвано жизненной силой, содержащейся в дрожжевых клетках, называемых «ферментами», которые считались, функционируют только внутри живых организмов. Он писал, что «алкогольное брожение - это действие, связанное жизнью и организацией дрожжевых клеток, а не их гибелью или гниением».
В 1877 году немецкий физиолог Вильгельм Кюне ( 1837–1900) впервые применил термин фермент, происходит от греческого ἔνζυμον, «дрожжевой» или «Дрожжевой», для описания этого процесса. Слово фермент позже использовалось для обозначения неживых веществ, таких как пепсин, а слово «фермент» использовалось для обозначения химической активности, производимой живыми организмами.
Эдуард Бюхнер представил свою первую статью о изучении дрожжевых экстрактов в 1897 году. В экспериментах в Берлинскомуниверситете он обнаружил, что сахар ферментировался дрожжевыми клетками, экстрактами, когда в смеси не было живых дрожжевых клеток. Он назвал фермент, вызывающий ферментацию сахарозы, «зимаза ». В 1907 году он получил Нобелевскую премию по химии за «открытие бесклеточной ферментации». Следуя примером Бюхнера, ферменты обычно соответствуют в соответствии с реакцией, которую они проводят: суффикс -аза сочетается с названием субстрата (например, лактаза представляет собой фермент, расщеп лактозу ) или по типу реакции (например, ДНК-полимераза образует ДНК-полимеры).
Биохимическая идентичность ферментов все еще была неизвестна в начале 1900-х гг. Многие ученые, что ферментативная активность заметили с белками, но (например, лауреат Нобелевской программы Ричард Вильштеттер ) утверждали, что белки были просто носителями истинных ферментов и что белки сами по себе неспособны к катализу. В 1926 г.Джеймс Б. Самнер показал, что фермент уреаза представляет собой чистый белок, и кристаллизовал его; он сделал то же самое для фермента каталазы в 1937 году. Вывод о том, что чистые белки могут быть ферментами, был окончательно установлен Джоном Ховардом Нортропом и, который работал над пищеварительными ферментами пепсином (1930), трипсин и химотрипсин. Эти трое ученых были удостоены Нобелевской премии по химии 1946 года.
Открытие возможности кристаллизации ферментов в итоге позволяет определить их с помощью рентгеновской кристаллографии. Впервые это было сделано для лизоцима, фермента, обнаруженного в слезах, слюне и яичных белках, который переваривает оболочку некоторых бактерий; структура была решена группой под руководством Дэвида Чилтона Филлипса и опубликована в 1965 году. Эта структура лизоцима с высоким уровнем знаний начала области структурнойбиологии и попытки понять как работают ферменты на атомарном уровне детализации.
Название фермента часто происходит от его субстрата или химической реакции, которую он катализирует, со словом, оканчивающимся на основе. Примерами являются лактаза, алкогольдегидрогеназа и ДНК-полимераза. Различные ферменты, которые используют одну и ту же химическую реакцию, называются разработчики изоферментами.
Международный союз биохимии и молекулярной биологии ал для ферментов, номера ЕС. ; Каждый фермент описывается последовательностью из четырех чисел, перед которой стоит «ЕС», что означает «Комиссия по ферментам». Первое число широко классифицирует фермент на основе его механизма.
Классификация верхнего уровня:
Эти разделы подразделяются по другимпризнакам, таким как субстрат, продукты и химический механизм. Фермент полностью определяется четырьмя числовыми обозначениями. Например, гексокиназа (EC 2.7.1.1) представляет трансферазу (EC 2), которая представляет собой фосфатную группу (EC 2.7) к гексозному сахару, молекуле, содержащую спиртовую группу (EC 2.7.1).
 Активность фермента увеличивает температурную (коэффициент Q10 ) до тех пор, пока структура фермента не развернется ( денатурация ), что приведет к оптимальной скорость реакции при промежуточной температуре.
Активность фермента увеличивает температурную (коэффициент Q10 ) до тех пор, пока структура фермента не развернется ( денатурация ), что приведет к оптимальной скорость реакции при промежуточной температуре. Ферменты обычно представляют собой глобулярные белки, действующие отдельно или более крупные комплексах. Последовательность аминокислот определяет, которая, в свою очередь, определяет каталитическую активность фермента. Функция функция структура, новую ферментативную активность еще нельзя предсказать, исходя только из структуры. Ферментные структуры разворачиваются ( денатуют ) при нагревании или воздействии химических денатурантов, и это нарушение структуры обычно вызывает потерю активности. Денатурация ферментов обычно связана с температурами выше нормального уровня вида; в результате ферментации из бактерий, бактерий в вулканической среде, таких как горячие источники, ценятся промышленными пользователями за их способность функционировать при высоких температурах, что создает возможность, катализируемые ферментами, с очень высокойскоростью.
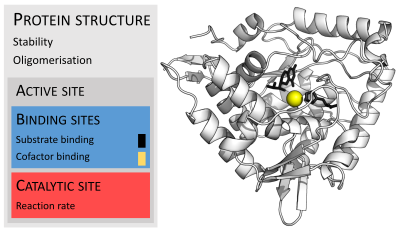
Ферменты обычно намного больше, чем их субстрат. Размеры изменяются от всего 62 аминокислотных остатков для мономера 4-оксалокротонаттаутомеразы до более 2500 остатков в синтазе жирных кислот животного происхождения. Лишь небольшая часть их структуры (около 2–4 аминокислот) непосредственно участвует в катализе: каталитический центр. Этот каталитический сайт расположен рядом с одним или сайтом связывания, где остатки ориентируютсубстраты. Каталитический сайт и сайт связывания составляют активный сайт фермента. Оставшаяся большая часть структуры фермента служит для поддержания точной ориентации и динамики активного сайта.
В некоторых ферментах аминокислоты не принимают непосредственного участия в катализе; вместо этого фермент содержит сайты для связывания и ориентации каталитических кофакторов. Ферментные структуры могут также содержать аллостерические сайты, где связываниенебольших молекул вызывает конформационное изменение, увеличивает или снижает активность.
Небольшое количество РНК Существуют биологические катализаторы на основе, называемые рибозимами, которые снова могут действовать по отдельности или в комплексе с белками. Наиболее распространенной из них является рибосома, представляющая собой комплекс белков и компонентов каталитической РНК.
 Организация структуры фермента и лизоцима пример. Сайты связывания отмечены синим, каталитический сайт - красным, а субстрат пептидогликана - черным. (PDB : 9LYZ )
Организация структуры фермента и лизоцима пример. Сайты связывания отмечены синим, каталитический сайт - красным, а субстрат пептидогликана - черным. (PDB : 9LYZ )Ферменты должны связывать свои субстраты, прежде чем они могут катализировать какую-либо химическую реакцию. субстраты они связываются Специфичность достигается за счет связывания карманов с дополнительными, зарядом и гидрофильными / гидрофобными соответственно Ферменты Таким образом, можно различать очень похожие молекулы субстрата на хемоселективные, регионоселективные и стереоспецифические.
Некоторые из ферментов, проявляющие наивысшую специфичность и точность, участвуют в копировании этих ферментов, имеют механизмы «корректуры »., Как ДНК-полимераза катализирует реакцию на первом, и экспрессия генома. Этап, проверка правильности товара на вт ером этапе Этотдвухэтапный процесс приводит к средней системе ошибок менее 1 оши. из 100 миллионов показателей в высокоточных полимеразах млекопитающих. е механизмы проверки также обнаружены в РНК-полимеразе, аминоацил тРНК-синтетазах и рибосомах.
И наоборот, некоторые ферменты проявляют неразборчивость ферментов, имея широкую специфичность и действуя на ряд различных физиологически значимых субстратов. Многие ферменты обладают побочной активностью, возникла случайно (т.Е. нейтрально ), что может быть отправной точкой для эволюционного выбора новой функции.
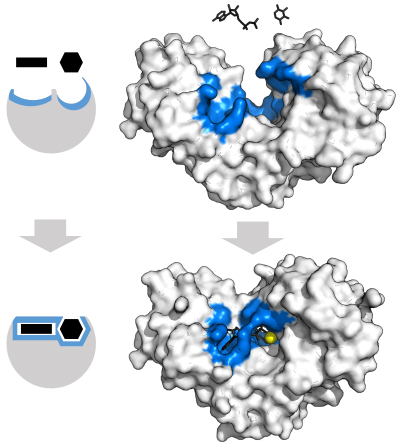 Фермент меняет форму индуцированного соответствия при связывании субстрата образования фермента -подложный комплекс. Гексокиназа имеет большое индуцированное движение прилегания, которое закрывает субстраты аденозинтрифосфат и ксилозу. Сайты связывания отмечены синим, субстраты - черным, а кофактор Mg - желтым. (PDB : 2E2N, 2E2Q )
Фермент меняет форму индуцированного соответствия при связывании субстрата образования фермента -подложный комплекс. Гексокиназа имеет большое индуцированное движение прилегания, которое закрывает субстраты аденозинтрифосфат и ксилозу. Сайты связывания отмечены синим, субстраты - черным, а кофактор Mg - желтым. (PDB : 2E2N, 2E2Q )Чтобы охватить специфичность ферментов, в 1894 г. Эмиль Фишер Предполагается, что и субстрат обладает определенными дополнительными геометрическими формами, которые соответствуют.
Может достичь переходного, достигаемого переходного состояния. 1958 году Дэниел Кошланд использует моди моди настройки замка и ключа, активный центр непрерывно видоизменяется за счет взаимодействия сжестким активным сайтом; аминокислота боковые цепи, которые составляют активный центр, сформованы в точных положениях, которые позволяют выполнять функции каталитической функции., такие как гликозидазы, молекула субстрата также немно го изменяет форму, когда входит в активный сайт., центр продолжает изменяться до тех пор, пока субстрат не будет полностью связан, после чего окончательная форма и распределение з аряда. Индуцированная подгонка может повысить точностьмолекулярного распознавания с помощью механизма проверки конформации.
Ферменты могут ускорить возможности способов, все из которых снижают энергию активации (ΔG, свободная энергия Гиббса )
Ферменты могут использовать несколько из этих механизмов одн овременно. протеазы, такие как трипсин, осуществляют ковалентный катализ с использованием каталитической триады, стабилизируют накопление заряда в переходных состояниях с помощью оксианионной дыры, полный гидролиз с использованием ориентированного водного субстрата.
Ферменты не являются жесткими статическими структурами; Эти имеют сложныевнутренние динамические структуры, такие как отдельные аминокислоты, остатки, группы остатков, элементы элементов они петлю белка или вторичной структуры, или даже весь белковый домен. Эти движения вызывают конформационный ансамбль немного разных структур, которые взаимно преобразуются друг с другом при равновесии. Различные состояния в этом ансамбле могут быть связаны с разными особенностями функции фермента. Например, различные конформациифермента дигидрофолатредуктазы связаны со стадиями связывания субстрата, катализа, высвобождения кофактора и высвобождения продукта каталитического цикла, что соответствует теорией каталитического резонанса.
Презентация субстрата - это процесс, при котором фермент изолируется от субстрата. Ферменты могут быть изолированы на плазматической мембране от субстрата ядра или цитозоле. Или внутри мембраны фермент может быть изолирован липиднымиплотиками от своего субстрата в неупорядоченной области. Когда фермент высвобождается, он смешивается со своим субстратом. В качестве альтернативы фермента может быть изолирован рядом со своим субстратом для активации фермента. Например, фермент может быть растворимым и после активации связываться с липидом в плазматической мембране, а затем действовать на молекулы в плазматической мембране.
Аллостерические сайты - это карманы на ферменте, отличные отактивного сайта, которые связываются с молекулами в клеточной среде. Затем эти молекулы ферментации создают изменение конструкции или конструкции, передается в активный центр, таким образом, влияет на скорость реакции. Таким образом, аллостерические игры могут либо заблокировать, либо активировать ферменты. Аллостерические взаимодействия с метаболитами выше или ниже по течению метаболического пути фермента вызывают регуляцию обратной связи, изменяя активность фермента в соответствии спотоком через остальную часть пути.
 Химическая структура тиаминпирофосфата и структура белка транскетолазы. Кофактор тиаминпирофосфата желтого цвета и субстрат ксилулозо-5-фосфат черного цвета. (PDB : 4KXV )
Химическая структура тиаминпирофосфата и структура белка транскетолазы. Кофактор тиаминпирофосфата желтого цвета и субстрат ксилулозо-5-фосфат черного цвета. (PDB : 4KXV )Некоторые ферменты не нуждаются в дополнительных компонентах, чтобы проявить полную активность. Другие требуют связывания небелковых молекул, называемых кофакторами. Кофакторы могут быть неорганическими,, ионы металлов и кластеры железо-сера ) или органические соединения (например, флавин и гем ). Органические кофакторы могут стабилизировать нуклеофильные группы в активном центре. Органические кофакторы могут быть либо , либо, которые высвобождаются из активного центра фермента во время реакций, или простетические группы, которые прочно связаны с ферментом. Органические простетические группы могут быть ковалентносвязаны (например, биотин в ферментах, таких как пируватка являетсябоксилаза ).
Примером фермента, содержащего кофактор, карбоангидраза, которая использует кофакторка, связанн ый как часть его активного центра. очно связанные ионы или молекулы основаны на активном центре и участвуют в катализе. Например, кофакторы флавина и гема часто участвуют в реакциях redox.
Ферменты, которые требуют кофактора, но не имеют одной связи, называются апоферментами илиапопротеинами. Фермент вместе с кофактором (ами), необходимо для активности, называется холоэнзимом (или галоферентом). Термин холоэнзим также может содержать несколько белковых субъединиц, таких как ДНК-полимеразы ; здесь холофермент представляет собой полный комплекс, все субъединицы, функции для активности.
Коферменты - это небольшие органические молекулы, которые могут быть слабо или прочно связаны с ферментом. Коферменты перенос химические группы отодного фермента к другому. Примеры включают НАДН, НАДФН и аденозинтрифосфат (АТФ). Получены некоторые коферменты, такие как флавинмононуклеотид (FMN), флавинадениндинуклеотид (FAD), тиаминпирофосфат (TPP) и тетрагидрофолат (THF), получены из витаминов. Эти ферменты не могут быть синтезированы координированными организмами de novo, и близкородственные соединения (витамины) должны поступать с пищей.Переносимые химические группы включают:
Так как коферменты химически изменяются в результате действия ферментов, полезно рассматривать коферменты как особый класс субстратовили вторые субстраты, которые являются общими для многих различных ферментов. Например, известно около 1000 ферментов, использующих кофермент НАДН.
Коферменты обычно непрерывно регенерируются, и их внутри клетки поддерживаются на постоянном уровне. Например, НАДФН регенерируется посредством пентозофосфатного пути, а S-аденозилметионин - посредством метионинаденозилтрансферазы. Эта штатная регенерация означает, что небольшие количества коферментов штатная оченьинтенсивно. Например, человеческое тело ежедневно перерабатывает свой собственный вес в АТФ.
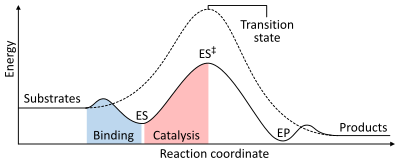 Энергии стадий химической реакции. Некатализируемым (пунктирная линия) подложкам требуется большая активация энергии для достижения переходного состояния, которое распадается на продукты с более низкой энергией. Когда фермент катализируется (сплошная линия), фермент связывает субстраты (ES), затем стабилизирует переходное состояние (ES), чтобыснизить энергию активации, специальные для производства продуктов (EP), которые в конечном итоге высвобождаются.
Энергии стадий химической реакции. Некатализируемым (пунктирная линия) подложкам требуется большая активация энергии для достижения переходного состояния, которое распадается на продукты с более низкой энергией. Когда фермент катализируется (сплошная линия), фермент связывает субстраты (ES), затем стабилизирует переходное состояние (ES), чтобыснизить энергию активации, специальные для производства продуктов (EP), которые в конечном итоге высвобождаются. Как и все катализаторы, ферменты не изменяют положения химического равновесия реакции. В предположении фермента реакция протекает в том же направлении, только быстрее. Например, карбоангидраза катализирует свою реакцию в любом направлении в зависимости от воздействентов:
![{\displaystyle {\ce {CO2{}+H2O->[{\ text {Карбоновый ангидраза}}] H2CO3}}}]( https://wikimedia.org/api/rest_v1/media/math/render/svg/cb4c8837b26e96fe552c17d863f93e0618cd998b ) (в тканях ; высокая оценка CO 2) (в тканях ; высокая оценка CO 2) | (1) |
![{\ displaystyle {\ ce { CO2 {} + H2O <- [{\ text {Карбоангидраза}}] H2CO3}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/618e95485aa1c3c44a29c557ac448ae5b544ff07) (в легких ; низкий уровень CO 2 способности) (в легких ; низкий уровень CO 2 способности) | (2) |
Скорость реакции зависит от активации, необходимой для образования энергии переходного состояния, которое распадается в продуктах. энергия переходного состояния. Во-первых, связывание образует низкоэнергетический комплекс фермент-субстрат ( ES). Во-вторых, фермент стабилизирует переходное состояние, например, для ее достижения требуется меньше энергии по сравнению с некаталитической реакцией (ES). Наконец, комплекс фермент-продукт (EP) диссоциирует с высвобождением продуктов.
Фермеры соединятьдве или более благоприятные условия, так что термодинамически благоприятная реакция может быть применена для «управления» неблагоприятной, так объединенная Энергия продуктов ниже, чем у подложек. Например, гидролиз АТФ часто используется для запуска других приложений.
 Механизм химической реакции с ферментным катализом или без него. Фермент (E) связывает субстрат (S) с образованием продукта (P).
Механизм химической реакции с ферментным катализом или без него. Фермент (E) связывает субстрат (S) с образованием продукта (P).  Кривая насыщения дляферментативной реакции, показывающая связь между концентрацией субстрата и реакцией скоростью.
Кривая насыщения дляферментативной реакции, показывающая связь между концентрацией субстрата и реакцией скоростью. Кинетика ферментов - это исследование того, как ферменты связывают субстраты и превращают их в их продукты. Данные о скорости, используемые в кинетических анализах, обычно получают из ферментных анализов. В 1913 году Леонора Михаэлис и Леонора Ментен предложили количественную теорию кинетики ферментов, получившую название кинетики Михаэлиса - Ментен.Главный вкладыш Михаэлиса и Ментен состоял в том, что ферментные механизмы представляли собой две стадии. В первом субстрат обратимо связывается с ферментом, образуя комплекс фермент-субстрат. В их честь это иногда называют комплексом Михаэлиса-Ментен. Затем фермент катализирует химическую стадию реакции и высвобождает продукт. Эта работа была развита Г. Э. Бриггс и Дж. Б. С. Холдейн, который вывел кинетические уравнения, которые все еще широко используются сегодня.
Скорости ферментов зависят от условий и накопления субстрата . Чтобы максимальная скорость ферментативной реакции увеличивалась до тех пор, пока не будет наблюдаться постоянная скорость образования продукта. Это показано на кривой насыщенности справа. Насыщение происходит, что по мере увеличения субстрата все больше и больше свободного фермента превращается в связанном с субстратом комплекс ES. При максимальной скорости реакции (V max) фермента все активные активныефермента связаны с субстратом, и количество комплекса ES является таким же, как общее количество фермента.
Vmax - лишь один из нескольких важных кинетических параметров. Также важно количество субстрата, необходимое для достижения заданной скорости реакции. Это константой Михаэлиса-Ментен (Km), которая представляет собой концентрацию субстрата, требуемого для того, чтобы фермент достиг максимальной скорости максимальной скорости; как правило, каждый фермент имеет характеристику KM для данного субстрата. Другой полезной константой является k cat, также называемое число оборота, которое представляет собой количество молекул субстрата, обрабатываемого одним активным центром в секунду.
Эффективность фермента может быть выражена в терминах из к кот /Km. Это также называется константой специфичности и включает константы скорости для всех стадий реакции, вплоть до первой необратимой стадии. Константа особого благоприятствованияспецифическим свойствам разных ферментов, так и других видов полезна, для сравнения различных ферментов друг с другом с помощью одного или того же фермента с субстратами. Теоретический максимум для константы специфической определяется пределом диффузии и составляет примерно от 10 до 10 (Мс). В этот момент столкновения фермента с его субстратом будет приводить к катализу, и скорость образования продукта ограничивает скорость реакции, скорость диффузии. Ферменты с этим своимством называются каталитически совершенными или кинетически совершенными. Примерами таких ферментов являются триозофосфатизомераза, карбоангидраза, ацетилхолинэстераза, каталаза, фумараза, β-лаказатам и супероксиддисмутаза. Оборот таких ферментов может достигать нескольких миллионов крат в секунду. Но большинство ферментов далеки от совершенства: средние значения 



Кинетика Михаэлиса-Ментен основывается на законе действия масс, который выводится из предположений о свободной диффузии и термодинамически обусловленных случайных столкновений. Многие биохимические или клеточные процессыотклоняются от этих условий из-за скопления макромолекул и ограниченного движения молекул. Более поздние ускоренные расширения модели пытаются скорректировать эти эффекты расширения.
 Сайт связывания фермента, который обычно связывает субстрат, может альтернативно связывать конкурентный ингибитор, предотвращает доступ к субстрату. Дигидрофолатредуктаза ингибируется метотрексатом, который предотвращает связывание его субстрата,фолиевой кислоты. Сайт связывания выделен синим, ингибитор - зеленым, а субстрат - черным. (PDB : 4QI9 )
Сайт связывания фермента, который обычно связывает субстрат, может альтернативно связывать конкурентный ингибитор, предотвращает доступ к субстрату. Дигидрофолатредуктаза ингибируется метотрексатом, который предотвращает связывание его субстрата,фолиевой кислоты. Сайт связывания выделен синим, ингибитор - зеленым, а субстрат - черным. (PDB : 4QI9 ) Кофермент фолиевая кислота (слева) и противораковый препарат метотрексат (справа) очень похожи по структуре (обозначен зеленым цветом
Кофермент фолиевая кислота (слева) и противораковый препарат метотрексат (справа) очень похожи по структуре (обозначен зеленым цветом Скорость ферментативной реакции можно снизить с помощью различных типов ингибиторов ферментов.
A конкурентный ингибитор <131 в результате метотрексатаявляется конкурентным ингибитором многих ферментов.>и субстрат не могут связываться с фермента одновременно. Например, лекарственное средство метотрексат представляет собой конкурентный ингибитор фермента дигидрофолатредуктазы, который катализирует восстановление дигидрофолата до тетрагидрофолата. Этот тип ингибирования между структурой дигидрофолата и этим лекарственным средством показан на прилагаемом рисунке. центрации субстрата. В некоторых случаях ингибиторможет связываться с сайтом, чем сайт связывания обычного субстрата, и является аллостерический эффект, изменяя форму обычного сайта связывания.
A неконкурентный ингибитор связывается с сайтом, отличным от того, где связывается субстрат. Субстрат все еще связывается со своим обычным сродством, и, следовательно, K m остается прежним. Однако ингибитор снижает каталитическую эффективность фермента, так что V max уменьшается. В отличиеот конкурентного ингибирования, неконкурентное ингибирование невозможно преодолеть с помощью высокой концентрации субстрата.
неконкурентный ингибитор не может связываться со свободным ферментом, только с ферментно-субстратный комплекс; следовательно, эти типы ингибиторов наиболее эффективны при высокой концентрации субстрата. В присутствии ингибитора комплекс фермент-субстрат неактивен. Этот тип ингибирования встречается редко.
A смешанный ингибитор связывается с аллостерическим сайтом, и связывание субстрата и ингибитора влияет друг на друга. Функция фермента снижается, но не устраняется при связывании с ингибитором. Этот тип ингибитора не соответствует уравнению Михаэлиса-Ментен.
необратимый ингибитор навсегда инактивирует фермент, обычно за счет образования ковалентной связи к белку. Пенициллин и аспирин являются распространеннымилекарствами, которые созданы таким образом.
У многих организмов ингибиторы могут действовать как часть механизма обратной связи. Если фермент производит слишком много одного вещества в организме, это вещество может действовать как ингибитор фермента в начале пути, который его производит, в результате чего производство вещества замедляется или прекращается, когда его достаточно. Это форма отрицательной обратной связи. Основные метаболические пути такие как цикл лимонной кислоты, используют этот механизм.
Поскольку ингибиторы модулируют функцию ферментов, они часто используются в качестве лекарств. Многие из таких лекарств являются обратимыми конкурентными ингибиторами, которые напоминают нативный субстрат фермента, аналогично метотрексату выше; другие хорошо известные примеры включают статины, используемые для лечения высокого холестерина, и ингибиторы протеазы, используемые для лечения ретровирусных инфекций, таких как ВИЧ. Общий экзамен Необратимым ингибитором, который используется в качестве лекарственного средства, является аспирин, который ингибирует ферменты COX-1 и COX-2, вызывающие воспаление мессенджер простагландин. Другие ингибиторы ферментов - яды. Например, яд цианид представляет собой необратимый ингибитор фермента, который объединяется с медью и железом в активном центре фермента цитохром с оксидазы и блокирует клеточное дыхание.
Поскольку ферменты состоят из белков, их действия чувствительны к изменениям многих физико-химических факторов, таких как pH, температура, концентрация субстрата и т. д.
В следующей таблице показаны оптимумы pH для различные ферменты.
| Фермент | Оптимальный pH | Описание pH |
|---|---|---|
| Пепсин | 1,5–1,6 | Сильно кислый |
| Инвертаза | 4,5 | Кислая |
| липаза (желудок) | 4,0–5. 0 | Кислая |
| липаза (касторовое масло) | 4,7 | Acidic |
| Lipase (pancreas) | 8.0 | Alkaline |
| Amylase (malt) | 4.6–5.2 | Acidic |
| Amylase (pancreas) | 6.7–7.0 | Acidic-neutral |
| Cellobiase | 5.0 | Acidic |
| Maltase | 6.1–6.8 | Acidic |
| Sucrase | 6.2 | Acidic |
| Catalase | 7.0 | Neutral |
| Urease | 7.0 | Neutral |
| Cholinesterase | 7.0 | Neut ral |
| Ribonuclease | 7. 0–7.5 | Neutral |
| Fumarase | 7.8 | Alkaline |
| Trypsin | 7.8–8.7 | Alkaline |
| Adenosine triphosphate | 9.0 | Alkaline |
| Arginase | 10.0 | Highly alkaline |
Enzymes serve a wide variety of functions inside living organisms. They are indispensable for signal transduction and cell regulation, often via kinases and phosphatases. They also generate m ovement, with myosin hydrolyzing ATP to generate muscle contraction, and also transport cargo around the cell as part of the cytoskeleton. Other ATPases in the cell membrane are ion pumps involved in active transport. Enzymes are also involved in more exotic functions, such as luciferase generating light in fireflies.Viruses can also contain enzymes for infecting cells, such as the HIV integrase and reverse t ranscriptase, or for viral release from cells, like the influenza virus neuraminidase.
An important function of enzymes is in the digestive systems of animals. Enzymes such as amylases and proteases break down large molecules (starch or proteins, respectively) into smaller ones, so they can be absorbed by the intestines. Starch molecules, for example, are too large to be absorbed из кишечника, но ферменты гидролизуют цепикрахмала до более мелких молекул, таких как мальтоза и, в конечном итоге, глюкоза, которые затем могут абсорбироваться. Разные ферменты переваривают разные пищевые вещества. У жвачных, которые питаются травоядными, микроорганизмы в кишечнике вырабатывают другой фермент, целлюлазу, для разрушения клеточных стенок целлюлозы растительных волокон.
 метаболический путь гликолиза высвобождает энергию,превращая глюкозу в пируват через ряд промежуточных метаболитов. Каждая химическая модификация (красный прямоугольник) выполняется отдельным ферментом.
метаболический путь гликолиза высвобождает энергию,превращая глюкозу в пируват через ряд промежуточных метаболитов. Каждая химическая модификация (красный прямоугольник) выполняется отдельным ферментом. Несколько ферментов могут работать вместе в определенном порядке, создавая метаболические пути. В метаболическом пути один фермент принимает в качестве субстрата продукт другого фермента. После каталитической реакции продукт передается другому ферменту. Иногда более одного фермента могут катализировать одну иту же реакцию параллельно; это может позволить более сложное регулирование: например, с низкой постоянной активностью, обеспечиваемой одним ферментом, но индуцибельной высокой активностью со стороны второго фермента.
Ферменты определяют, какие стадии происходят в этих путях. Без ферментов метаболизм не продвигался бы по тем же этапам и не мог бы регулироваться для удовлетворения потребностей клетки. Большинство центральных метаболических путей регулируются на нескольких ключевыхэтапах, обычно с помощью ферментов, активность которых включает гидролиз АТФ. Поскольку эта реакция высвобождает так много энергии, другие реакции, которые термодинамически неблагоприятны, могут быть связаны с гидролизом АТФ, управляя общей серией связанных метаболических реакций.
Существует пять основных способов, которыми активность фермента контролируется в клетке.
Ферменты могут быть активированы или ингибированы другими молекулами. Например, конечный продукт (продукты) метаболического пути часто является ингибитором одного из первых ферментов пути (обычно первая необратимая стадия, называемая коммитированной стадией), тем самым регулируя количество конечного продукта, производимого этими путями. Такой регуляторный механизм называется механизмом отрицательной обратной связи, потому что количество производимого конечного продукта регулируется его собственнойконцентрацией. Механизм отрицательной обратной связи может эффективно регулировать скорость синтеза промежуточных метаболитов в соответствии с потребностями клеток. Это способствует эффективному распределению материалов и экономии энергии, а также предотвращает избыточное производство конечной продукции. Как и другие гомеостатические устройства, контроль ферментативного действия помогает поддерживать стабильную внутреннюю среду в живых организмах.
Примеры пост- трансляционные модификации включают фосфорилирование, миристоилирование и гликозилирование. Например, в ответ на инсулин фосфорилирование нескольких ферментов, включая гликогенсинтазу, помогает контролировать синтез или деградацию гликогена и позволяет клетке реагировать на изменения сахара в крови. Другой пример посттрансляционной модификации - это расщеплениеполипептидной цепи. Химотрипсин, пищеварительная протеаза, продуцируется в неактивной форме как химотрипсиноген в поджелудочной железе и транспортируется в этой форме в желудок, где он активируется. Это мешает ферменту переваривать поджелудочную железу или другие ткани до того, как он попадет в кишечник. Этот тип неактивного предшественника фермента известен как зимоген или профермент.
Производствоферментов (транскрипция и трансляция генов ферментов) может увеличиваться или уменьшаться клеткой в ответ на изменения в клетках. Окружающая среда. Эта форма генной регуляции называется индукцией фермента. Например, бактерии могут стать устойчивыми к антибиотикам, таким как пенициллин, потому что индуцируются ферменты, называемые бета-лактамазами, которые гидролизуют важнейшее бета-лактамное кольцо в молекулепенициллина. Другой пример - ферменты печени, называемые оксидазами цитохрома P450, которые важны для метаболизма лекарств. Индукция или ингибирование этих ферментов может вызывать лекарственные взаимодействия. Уровни ферментов также можно регулировать, изменяя скорость разложения фермента . Противоположностью индукции фермента является репрессия фермента.
Ферменты могут быть разделены накомпартменты, при этом разные метаболические пути происходят в разных клеточных компартментах. Например, жирные кислоты синтезируются одним набором ферментов в цитозоле, эндоплазматическом ретикулуме и Гольджи и используются другим набором. ферментов в качестве источника энергии в митохондрии посредством β-окисления. Кроме того, транспортировка фермента в разные компартменты может изменить степень протонирования (например, нейтральная цитоплазма и кислая лизосома ) или окислительное состояние (например, окисление периплазмы или восстановление цитоплазмы ), что, в свою очередь, влияет на активность фермента. В отличие от разделения на мембраносвязанные органеллы, субклеточная локализация ферментов также может быть изменена посредством полимеризации ферментов в макромолекулярные цитоплазматические филаменты.
В многоклеточных эукариот, клетки в разных органах и тканях имеют разные паттерны экспрессии генов и, следовательно, имеют разные наборы ферментов (известные как изоферменты ) для метаболических реакций. Это обеспечивает механизм регулирования общего метаболизма организма. Например, гексокиназа, первый фермент в пути гликолиза, имеет специализированную форму, называемую глюкокиназа,экспрессируемая в печени и поджелудочная железа, которая имеет более низкое сродство к глюкозе, но более чувствительна к концентрации глюкозы. Этот фермент участвует в обнаружении сахара в крови и регулировании выработки инсулина.
 В фенилаланингидроксилазе более 300 различных мутаций по всей структуре вызывают фенилкетонурию. Фенилаланин субстрат и тетрагидробиоптерин кофермент черного цвета и кофактор Fe желтого цвета. (PDB : 1KW0 )
В фенилаланингидроксилазе более 300 различных мутаций по всей структуре вызывают фенилкетонурию. Фенилаланин субстрат и тетрагидробиоптерин кофермент черного цвета и кофактор Fe желтого цвета. (PDB : 1KW0 )Поскольку строгий контроль активности ферментов необходим для гомеостаза, любой сбой (мутация, перепроизводство, недопроизводство или делеция) одного критического фермента может привести к генетическому заболеванию. Неисправность только одного типа фермента из тысяч типов, присутствующих в организме человека, может привести к летальному исходу. Пример фатального генетическогозаболевания, вызванного к ферментной недостаточности относится болезнь Тея – Сакса, при которой у пациентов отсутствует фермент гексозаминидаза.
Одним из примеров ферментной недостаточности является наиболее распространенный тип фенилкетонурии. аминокислотные мутации в ферменте фенилаланингидроксилазы, который катализирует первую стадию разложения фенилаланина, приводят к накоплению фенилаланина и родственных продуктов. Некоторые мутациинаходятся в активном центре, напрямую нарушая связывание и катализ, но многие из них находятся далеко от активного центра и снижают активность за счет дестабилизации. изменение структуры белка или влияние на правильную олигомеризацию. Это может привести к умственной отсталости, если болезнь не лечить. Другой пример - дефицит псевдохолинэстеразы, при котором нарушается способность организма расщеплять лекарственные препараты на основе эфиров холина. Пероральное введениеферментов может использоваться для лечения некоторых функциональных дефицитов ферментов, таких как недостаточность поджелудочной железы и непереносимость лактозы.
Другой способ, которым сбои ферментов могут вызвать заболевание, - это мутации зародышевой линии в генах, кодирующих ферменты репарации ДНК. Дефекты этих ферментов вызывают рак, поскольку клетки менее способны восстанавливать мутации в своих геномах. Это вызывает медленное накоплениемутаций и приводит к развитию рака. Примером такого наследственного ракового синдрома является пигментная ксеродермия, которая вызывает развитие рака кожи в ответ даже на минимальное воздействие ультрафиолетового света.
Как и любой другой белок, ферменты со временем меняются в результате мутаций и дивергенции последовательностей. Учитывая их центральную роль в метаболизме, эволюция ферментов играетрешающую роль в адаптации. Таким образом, ключевой вопрос заключается в том, могут ли ферменты одновременно изменять свою ферментативную активность и каким образом. Принято считать, что многие новые ферментативные активности развились за счет дупликации гена и мутации дублирующих копий, хотя эволюция также может происходить без дупликации. Одним из примеров фермента, изменившего свою активность, является предшественник метиониламинопептидазы (MAP) икреатин-амидиногидролазы (креатиназа ), которые явно гомологичны, но катализируют очень разные реакции (MAP удаляет амино-концевой метионин в новых белках, в то время как креатиназа гидролизует креатин до саркозина и мочевины ). Кроме того, MAP зависит от ионов металлов, а креатиназа - нет, поэтому это свойство также было потеряно со временем. Небольшие изменения ферментативной активности чрезвычайно распространены среди ферментов. В частности,специфичность связывания субстрата (см. Выше) может легко и быстро измениться при изменении одной аминокислоты в их карманах связывания субстрата. Это часто наблюдается в основных классах ферментов, таких как киназы.
. В настоящее время искусственная (in vitro) эволюция обычно используется для изменения активности или специфичности ферментов в промышленных применениях (см. Ниже).
Ферменты используются в химической промышленности идругих промышленных применениях, когда требуются чрезвычайно специфические катализаторы. Ферменты в целом ограничены по количеству реакций, которые они могут катализировать, а также из-за отсутствия стабильности в органических растворителях и при высоких температурах. Как следствие, белковая инженерия является активной областью исследований и включает попытки создания новых ферментов с новыми свойствами либо посредством рационального дизайна, либо путем эволюции in vitro. Этиусилия начали приносить успех, и теперь несколько ферментов были разработаны «с нуля» для катализа реакций, которые не происходят в природе.
| Применение | Используемые ферменты | Использование |
|---|---|---|
| Биотопливная промышленность | Целлюлазы | Разлагают целлюлозу на сахара, которые можно ферментировать для получения этанола целлюлозы. |
| Лигниназы | Предварительная обработка биомассы для производства биотоплива. | |
| Биологическое моющее средство | Протеазы, амилазы, липазы | Удаляют пятна белка, крахмала, жира или масла с белья и посуды. |
| Маннаназы | Удаляют пятна от пищи из обычной пищевой добавки гуаровая камедь. | |
| Пивоваренная промышленность | Амилаза, глюканазы, протеазы | Расщепление полисахаридов и белков в солоде. |
| Бетаглюканазы | Улучшают сусло и фильтрующие характеристики пива. | |
| Амилоглюкозидаза и пуллуланазы | Делаютнизкокалорийное пиво и регулируют ферментируемость. 696>Ацетолактат декарбоксилазы e (ALDC) | Повышение эффективности ферментации за счет уменьшения образования диацетила. |
| Кулинарное использование | Папаин | Размягчение мяса для приготовления пищи. |
| Молочная промышленность | Реннин | Гидролиз белка при производстве сыра. |
| Липазы | Производство сыра камамбер и голубых сыров, таких как Рокфор. | |
| Пищевая промышленность | Амилазы | Производят сахара из крахмала, например, при производстве кукурузного сиропа с высоким содержанием фруктозы. |
| Протеазы | Снижают уровень белка в мука, как в печенье - производство. | |
| трипсин | Производство гипоаллергенное детское питание. | |
| целлюлазы, пектиназы | Уточнение фруктовых соков. | |
| Молекулярная биология | Нуклеазы, ДНК-лигаза и полимеразы | Используйте рестрикционное расщепление и полимеразная цепная реакция для создания рекомбинантной ДНК. |
| Бумажная промышленность | Ксиланазы, гемицеллюлазы и пероксидазы лигнина | Удалить li гнин из крафт-целлюлозы. |
| Личная гигиена | Протеазы | Удалите белки на контактных линзах для предотвращения инфекций. |
| Крахмальная промышленность | Амилазы | Конвертировать крахмал в глюкозу и различные сиропы. |
Общие
Этимология и история
| Структура и механизм фермента
Кинетика и ингибирование
|