
Войти
| Найдите аллостерическую в Wiktionary, бесплатном словарь. |
 Аллостерическая регуляция фермента
Аллостерическая регуляция фермента В биохимии, аллостерическая регуляция (или аллостерическая регуляция ) - это регуляция фермента путем связывания эффекторной молекулы с сайтом, отличным от активного сайта фермента.
Сайт, с которым связывается эффектор, называется аллостерическим сайтом или регуляторным сайтом. Аллостерические сайты позволяют эффекторам связываться с белком, что часто приводит к конформационным изменениям, связанным с динамикой белка. Эффекторы, которые усиливают активность белка, называются аллостерическими активаторами, тогда как те, которые снижают активность белка, называются аллостерическими ингибиторами.
Аллостерические правила являются естественным примером контуров управления, таких как обратная связь от последующих продуктов или прямая связь с исходных субстратов. Аллостерия на большие расстояния особенно важна в передаче сигналов клеток. Аллостерическая регуляция также особенно важна для способности клетки регулировать активность фермента.
Термин аллостерия происходит от древнегреческого allos (ἄλλος ), «другой» и стереосигнала (στερεὀς ), «твердый ( объект) ". Это относится к тому факту, что регуляторный сайт аллостерического белка физически отличается от его активного сайта.
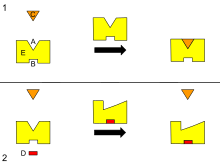 A - Активный сайт. B - Аллостерический сайт. C - Субстрат. D - Ингибитор. E - Фермент. Это диаграмма аллостерической регуляции фермента.
A - Активный сайт. B - Аллостерический сайт. C - Субстрат. D - Ингибитор. E - Фермент. Это диаграмма аллостерической регуляции фермента. Многие аллостерические эффекты могут быть объяснены согласованной моделью MWC, предложенной Монод, Вайманом и Changeux, или последовательной моделью, описанной Кошландом, Немети, и Филмер. Оба постулируют, что белковые субъединицы существуют в одной из двух конформаций, напряженной (Т) или расслабленной (R), и что расслабленные субъединицы связываются с субстратом легче, чем субъединицы в напряженном состоянии. Две модели больше всего различаются своими предположениями о взаимодействии субъединиц и предсуществовании обоих состояний. Для белков, в которых субъединицы существуют более чем в двух конформациях, может использоваться ландшафтная модель аллостерии, описанная Куэндет, Вайнштейном и ЛеВином.
Согласованная модель аллостерии, также называемая моделью симметрии или моделью MWC, постулирует, что субъединицы фермента связаны таким образом, что конформационная изменение в одной субъединице обязательно передается всем другим субъединицам. Таким образом, все субъединицы должны существовать в одной и той же конформации. Модель также утверждает, что в отсутствие какого-либо лиганда (субстрата или другого) равновесие благоприятствует одному из конформационных состояний, T или R. Равновесие может быть смещено в состояние R или T посредством связывания одного лиганд (аллостерический эффектор или лиганд) к сайту, который отличается от активного сайта (аллостерический сайт).
Последовательная модель аллостерической регуляции утверждает, что субъединицы не связаны таким образом, что конформационное изменение в одной вызывает аналогичное изменение в других. Таким образом, все субъединицы фермента не требуют одинаковой конформации. Более того, последовательная модель диктует, что молекулы субстрата связываются посредством протокола индуцированной подгонки. Хотя такая индуцированная подгонка переводит субъединицу из напряженного состояния в расслабленное состояние, она не распространяет конформационные изменения на соседние субъединицы. Вместо этого связывание с субстратом одной субъединицы лишь незначительно изменяет структуру других субъединиц, так что их сайты связывания становятся более восприимчивыми к субстрату. Подводя итог:
Модель аллостерической регуляции морфеина - это диссоциативная согласованная модель.
Морфеин - это гомоолигомерная структура, которая может существовать как ансамбль физиологически значимых и функционально различных альтернативных четвертичных сборки. Переходы между альтернативными сборками морфеина включают диссоциацию олигомеров, конформационные изменения в диссоциированном состоянии и повторную сборку с образованием другого олигомера. Требуемый этап разборки олигомера отличает модель морфеина для аллостерической регуляции от классических моделей MWC и KNF.
Порфобилиногенсинтаза (PBGS) является прототипом морфеина.
Ансамблевые модели аллостерической регуляции перечисляют статистический ансамбль аллостерической системы как функцию ее функции потенциальной энергии, а затем связывают конкретные статистические измерения аллостерии к определенным энергетическим элементам в энергетической функции (например, межмолекулярный солевой мостик между двумя доменами). Ансамблевые модели, такие как ансамблевая аллостерическая модель и аллостерическая модель Изинга, предполагают, что каждая область системы может принимать два состояния, аналогичные модели MWC. Модель ландшафта аллостерии, введенная Куэндет, Вайнштейном и ЛеВином, позволяет доменам иметь любое количество состояний, и вклад определенного молекулярного взаимодействия в данное аллостерическое взаимодействие можно оценить с помощью строгого набора правил. Молекулярная динамика моделирование можно использовать для оценки статистического ансамбля системы, чтобы его можно было проанализировать с помощью ландшафтной модели аллостерии.
Аллостерическая модуляция используется для изменения активности молекул и ферментов в биохимии и фармакологии. Для сравнения, типичное лекарство создается для связывания с активным сайтом фермента, что, таким образом, запрещает связывание субстрата с этим ферментом, вызывая снижение активности фермента. Аллостерическая модуляция происходит, когда эффектор связывается с аллостерическим сайтом (также известным как регуляторный сайт) фермента и изменяет активность фермента. Аллостерические модуляторы предназначены для того, чтобы соответствовать аллостерическому сайту, чтобы вызвать конформационное изменение фермента, в частности изменение формы активного сайта, которое затем вызывает изменение его активности. В отличие от обычных лекарств модуляторы не являются конкурентными ингибиторами. Они могут быть положительными (активирующими), вызывающими повышение активности фермента, или отрицательными (ингибирующими), вызывающими снижение активности фермента. Использование аллостерической модуляции позволяет контролировать эффекты специфической активности ферментов; в результате аллостерические модуляторы очень эффективны в фармакологии. В биологической системе может быть трудно отличить аллостерическую модуляцию от модуляции с помощью презентации субстрата.
Пример этой модели можно увидеть с Mycobacterium tuberculosis, бактерия, которая идеально приспособлена к жизни в макрофагах человека. Сайты фермента служат связью между различными субстратами. В частности, между AMP и G6P. Такие сайты также служат в качестве механизма определения эффективности фермента.
Положительная аллостерическая модуляция (также известная как аллостерическая активация) происходит, когда связывание одного лиганда усиливает притяжение между молекулами субстрата и другими сайтами связывания. Примером является связывание молекул кислорода с гемоглобином, где кислород эффективно является как субстратом, так и эффектором. Аллостерический или «другой» сайт представляет собой активный сайт соседней белковой субъединицы. Связывание кислорода с одной субъединицей вызывает конформационные изменения в этой субъединице, которая взаимодействует с оставшимися активными центрами, повышая их сродство к кислороду. Другой пример аллостерической активации наблюдается в цитозольной IMP-GMP специфической 5'-нуклеотидазе II (cN-II), где сродство к субстрату GMP увеличивается при связывании GTP на границе раздела димеров.
Отрицательная аллостерическая модуляция (также известная как аллостерическое ингибирование) происходит, когда связывание одного лиганда снижает сродство к субстрату на других активных сайтах. Например, когда 2,3-BPG связывается с аллостерическим сайтом на гемоглобине, сродство к кислороду всех субъединиц снижается. Это когда в сайте связывания отсутствует регулятор.
Прямые ингибиторы тромбина представляют собой отличный пример отрицательной аллостерической модуляции. Обнаружены аллостерические ингибиторы тромбина, которые потенциально могут использоваться в качестве антикоагулянтов.
Другим примером является стрихнин, судорожный яд, который действует как аллостерический ингибитор рецептора глицина. Глицин является основным пост- синаптическим ингибирующим нейромедиатором в млекопитающих спинном мозге и стволе мозга. Стрихнин действует на отдельном сайте связывания на рецепторе глицина аллостерическим образом; т.е. его связывание снижает сродство рецептора глицина к глицину. Таким образом, стрихнин подавляет действие тормозящего медиатора, что приводит к судорогам.
Другой случай, в котором можно увидеть отрицательную аллостерическую модуляцию, - это между АТФ и ферментом фосфофруктокиназой в петле отрицательной обратной связи, которая регулирует гликолиз. Фосфофруктокиназа (обычно обозначаемая как PFK ) представляет собой фермент, который катализирует третью стадию гликолиза: фосфорилирование фруктозо-6-фосфата в фруктозу. 1,6-бисфосфат. PFK может аллостерически подавляться высокими уровнями АТФ в клетке. Когда уровни АТФ высоки, АТФ будет связываться с аллосторным участком на фосфофруктокиназе, вызывая изменение трехмерной формы фермента. Это изменение вызывает снижение его сродства к субстрату (фруктозо-6-фосфат и АТФ ) в активном центре, и фермент считается неактивным. Это вызывает прекращение гликолиза при высоком уровне АТФ, таким образом сохраняя глюкозу в организме и поддерживая сбалансированные уровни клеточного АТФ. Таким образом, АТФ служит отрицательным аллостерическим модулятором для PFK, несмотря на то, что он также является субстратом фермента.
Гомотропный аллостерический модулятор представляет собой субстрат для своего целевого фермента, а также регуляторную молекулу активности фермента. Обычно это активатор фермента. Например, O 2 и CO являются гомотропными аллостерическими модуляторами гемоглобина. Аналогичным образом, в IMP / GMP-специфической 5'-нуклеотидазе связывание одной молекулы GMP с одной субъединицей тетрамерного фермента приводит к увеличению сродства к GMP последующих субъединиц, как показывают графики зависимости сигмоидального субстрата от скорости.
Гетеротропный аллостерический модулятор - это регуляторная молекула, которая не является субстратом фермента. Это может быть активатор или ингибитор фермента. Например, H, CO 2 и 2,3-бисфосфоглицерат являются гетеротропными аллостерическими модуляторами гемоглобина. И снова в IMP / GMP-специфической 5'-нуклеотидазе связывание молекулы GTP на границе димера в тетрамерном ферменте приводит к увеличению сродства к субстрату GMP в активном центре, что указывает на гетеротропную аллостерическую активацию K-типа.
Как было подробно подчеркнуто выше, некоторые аллостерические белки могут регулироваться как их субстратами, так и другими молекулами. Такие белки способны к как гомотропным, так и гетеротропным взаимодействиям.
Некоторые аллостерические активаторы называют «основными» или «облигатными» активаторами в том смысле, что в их отсутствие, активность их целевого фермента очень низкая или пренебрежимо мала, как, например, в случае с активностью N-ацетилглутамата в отношении карбамоилфосфатсинтетазы I.
Не- регуляторный аллостерический сайт - это любой нерегулирующий компонент фермента (или любого белка), который сам не является аминокислотой. Например, многие ферменты требуют связывания натрия для обеспечения правильного функционирования. Однако натрий не обязательно действует как регуляторная субъединица; натрий присутствует всегда, и нет известных биологических способов добавления / удаления натрия для регулирования активности ферментов. Нерегулирующая аллостерия может включать любые другие ионы, помимо натрия (кальция, магния, цинка), а также другие химические вещества и, возможно, витамины.
Аллостерическая модуляция рецептора возникает в результате связывания аллостерических модуляторов в другом сайте («регуляторный сайт ») по сравнению с эндогенным сайтом. лиганд («активный сайт ») и усиливает или ингибирует эффекты эндогенного лиганда. В нормальных условиях он действует, вызывая конформационное изменение в молекуле рецептора, что приводит к изменению аффинности связывания лиганда. Таким образом, аллостерический лиганд модулирует активацию рецептора своим первичным ортостерическим лигандом, и можно думать, что он действует как диммер в электрической цепи, регулируя интенсивность ответа.
Например, ГАМК A рецептор имеет два активных сайта, которые связывает нейромедиатор гамма-аминомасляная кислота (ГАМК), но также имеет бензодиазепин и общий анестетик регуляторные сайты связывания. Каждый из этих регуляторных сайтов может вызывать положительную аллостерическую модуляцию, усиливая активность ГАМК. Диазепам является положительным аллостерическим модулятором в регуляторном сайте бензодиазепина, а его антидот флумазенил является антагонистом.
Более свежие примеры лекарств, которые аллостерически модулировать их цели, включая имитирующие кальций цинакальцет и лечение ВИЧ маравирок.
Аллостерические сайты могут представлять новую мишень для лекарств. Существует ряд преимуществ использования аллостерических модуляторов в качестве предпочтительных терапевтических агентов по сравнению с классическими ортостерическими лигандами. Например, сайты аллостерического связывания G-белкового рецептора (GPCR) не столкнулись с таким же эволюционным давлением, как ортостерические сайты, чтобы приспособиться к эндогенному лиганду, поэтому они более разнообразны. Следовательно, большая селективность GPCR может быть получена путем нацеливания на аллостерические сайты. Это особенно полезно для GPCR, где селективная ортостерическая терапия была затруднена из-за сохранения последовательности ортостерического сайта у подтипов рецепторов. Кроме того, эти модуляторы обладают пониженным потенциалом токсических эффектов, поскольку модуляторы с ограниченным взаимодействием будут иметь максимальный уровень своего действия, независимо от введенной дозы. Другой тип фармакологической селективности, который является уникальным для аллостерических модуляторов, основан на кооперативности. Аллостерический модулятор может проявлять нейтральную кооперативность с ортостерическим лигандом на всех подтипах данного рецептора, кроме интересующего подтипа, который называется «абсолютной селективностью подтипа». Если аллостерический модулятор не обладает заметной эффективностью, он может обеспечить другое мощное терапевтическое преимущество по сравнению с ортостерическими лигандами, а именно способность избирательно настраивать повышающие или понижающие тканевые ответы только при наличии эндогенного агониста. Олигомер-специфичные сайты связывания малых молекул являются лекарственными мишенями для медицинских морфеинов.
Существует множество синтетических соединений, содержащих несколько нековалентных сайтов связывания, которые демонстрируют конформационные изменения при занятии одного участка. Кооперативность между вкладами единичного связывания в таких супрамолекулярных системах является положительной, если занятие одного сайта связывания увеличивает аффинность ΔG во втором сайте, и отрицательной, если аффинность не выше. Большинство синтетических аллостерических комплексов основаны на конформационной реорганизации при связывании одного эффекторного лиганда, что затем приводит либо к усилению, либо к ослаблению ассоциации второго лиганда с другим сайтом связывания. Конформационное связывание между несколькими сайтами связывания в искусственных системах обычно намного больше, чем в белках с их обычно большей гибкостью. Параметр, определяющий эффективность (измеряемый соотношением констант равновесия Krel = KA (E) / KA в присутствии и отсутствии эффектора E), представляет собой конформационную энергию, необходимую для принятия закрытой или напряженной конформации для связывания лиганда. A.
Во многих поливалентных супрамолекулярных системах может происходить прямое взаимодействие между связанными лигандами, что может приводить к большому взаимодействию. Наиболее распространено такое прямое взаимодействие между ионами в рецепторах ионных пар. Это сотрудничество часто также называют аллостерией, хотя конформационные изменения здесь не обязательно запускают события связывания.
Аллостерия - это прямое и эффективное средство регулирования функции биологических макромолекул, производимое связыванием лиганда на аллостерическом сайте, топографически отличном от ортостерический узел. Ожидается, что из-за часто высокой селективности рецепторов и более низкой токсичности в отношении мишеней аллостерическая регуляция будет играть все большую роль в открытии лекарств и биоинженерии. База данных AlloSteric (ASD, http://mdl.shsmu.edu.cn/ASD ) обеспечивает центральный ресурс для отображения, поиска и анализа структуры, функции и связанных аннотаций для аллостерических молекул. В настоящее время ASD содержит аллостерические белки более 100 видов и модуляторы трех категорий (активаторы, ингибиторы и регуляторы). Каждый белок снабжен аннотацией с подробным описанием аллостерии, биологического процесса и связанных заболеваний, а каждый модулятор - сродством связывания, физико-химическими свойствами и терапевтической областью. Интеграция информации об аллостерических белках в РАС должна позволить предсказывать аллостерию для неизвестных белков с последующей экспериментальной проверкой. Кроме того, модуляторы, курируемые в ASD, могут использоваться для исследования потенциальных аллостерических мишеней для запрашиваемого соединения и могут помочь химикам реализовать модификации структуры для создания новых аллостерических лекарств.
Не все белковые остатки играют одинаково важные роли в аллостерической регуляции. Идентификация остатков, необходимых для аллостерии (так называемых «аллостерических остатков»), была в центре внимания многих исследований, особенно в последнее десятилетие. Частично этот растущий интерес является результатом их общей важности для науки о белках, но также и потому, что аллостерические остатки могут использоваться в биомедицинских контекстах. Фармакологически важные белки с сайтами, на которые трудно воздействовать, могут уступить место подходам, в которых один альтернативно нацелен на более легкие для доступа остатки, которые способны аллостерически регулировать первичный интересующий сайт. Эти остатки в целом можно классифицировать как поверхностные и внутренние аллостерические аминокислоты. Аллостерические сайты на поверхности обычно играют регулирующую роль, фундаментально отличную от таковых внутри; поверхностные остатки могут служить рецепторами или эффекторными сайтами в передаче аллостерических сигналов, тогда как те, что внутри могут действовать, чтобы передавать такие сигналы.