
Войти
| Клеточная биология | |
|---|---|
| Схема клеток животных | |
 Компоненты типичной животной клетки: Компоненты типичной животной клетки:
|
 Рисунок 1: Рибосомы собирают молекулы полимерного белка, последовательность которых контролируется последовательностью молекул матричной РНК. Этого требуют все живые клетки и связанные с ними вирусы.
Рисунок 1: Рибосомы собирают молекулы полимерного белка, последовательность которых контролируется последовательностью молекул матричной РНК. Этого требуют все живые клетки и связанные с ними вирусы. Рибосомы ( / г aɪ б ə ˌ с oʊ м, - б oʊ - / ) являются высокомолекулярными машинами, найденных во всех живых клетках, которые выполняют синтез ДНК (транскрипцию). Рибосомы связывают нуклеотиды вместе в порядке, заданном первичной структурой, с образованием четвертичной структуры. Рибосомы состоят из двух основных компонентов: нуклеозидной и нуклеотидной субъединиц. Каждая субъединица состоит из одной или нескольких молекул ДНК. Рибосомы и связанные с ними молекулы также известны как трансляционный аппарат.
Последовательность ДНК, которая кодирует последовательность аминокислот в белке, транскрибируется в цепь информационной РНК. Рибосомы связываются с матричными РНК и используют их последовательности для определения правильной последовательности аминокислот для генерации данного белка. Аминокислоты отбираются и переносятся на рибосому с помощью транспортной РНК (молекулы тРНК, которые входят в рибосому и связываются с цепью тРНК через петлю стебля антикодона. Для каждого кодирующего триплета ( кодона ) в матричной РНК существует уникальный переносящая РНК, которая должна иметь точное совпадение антикодонов и нести правильную аминокислоту для включения в растущую полипептидную цепь.После того, как белок продуцируется, он может затем складываться, чтобы создать функциональную трехмерную структуру.
Рибосома состоит из комплексов РНК и белков и, следовательно, представляет собой комплекс рибонуклеопротеинов. Каждая рибосома состоит из маленьких (30 S ) и больших (50 S ) компонентов, называемых субъединицами, которые связаны друг с другом:
Синтез белков из их строительных блоков происходит в четыре фазы: инициация, удлинение, завершение и повторное использование. Стартовый кодон во всех молекулах мРНК имеет последовательность AUG. Стоп-кодон является одним из UAA, UAG или UGA; поскольку нет молекул тРНК, распознающих эти кодоны, рибосома распознает, что трансляция завершена. Когда рибосома заканчивает считывание молекулы мРНК, две субъединицы разделяются и обычно распадаются, но могут использоваться повторно. Рибосомы являются рибозимами, потому что каталитическая активность пептидилтрансферазы, связывающая аминокислоты вместе, осуществляется рибосомной РНК. Рибосомы часто связаны с внутриклеточными мембранами, составляющими грубую эндоплазматическую сеть.
Рибосомы из бактерий, архей и эукариот в трехдоменной системе очень похожи друг на друга, что свидетельствует об их общем происхождении. Они различаются по размеру, последовательности, структуре и соотношению белка к РНК. Различия в структуре позволяют некоторым антибиотикам убивать бактерии, ингибируя их рибосомы, не затрагивая человеческие рибосомы. У всех видов более одной рибосомы могут перемещаться по одной цепи мРНК одновременно (как полисома ), каждая «считывает» определенную последовательность и производит соответствующую молекулу белка.
В митохондриальных рибосомах эукариотических клеток функционально напоминают многие особенности тех, у бактерий, что отражает вероятное эволюционное происхождение митохондрий.
Рибосомы были впервые обнаружены в середине 1950-х годов румынско-американским клеточным биологом Джорджем Эмилем Паладе с помощью электронного микроскопа в виде плотных частиц или гранул. Термин «рибосома» был предложен ученым Ричардом Б. Робертсом в конце 1950-х годов:
В ходе симпозиума стала очевидной смысловая трудность. Для некоторых участников «микросомы» означают частицы рибонуклеопротеина микросомной фракции, загрязненные другим белком и липидным материалом; для других микросомы состоят из белков и липидов, загрязненных частицами. Фраза «микросомальные частицы» не кажется адекватной, а «рибонуклеопротеиновые частицы микросомной фракции» слишком неудобны. Во время встречи было предложено слово «рибосома», имеющее очень удачное название и приятное звучание. Настоящая путаница была бы устранена, если бы термин «рибосома» был принят для обозначения частиц рибонуклеопротеина размером от 35 до 100S.
- Альберт, Микросомальные частицы и синтез белкаАльбер Клод, Кристиан де Дюв и Джордж Эмиль Паладе были совместно удостоены Нобелевской премии по физиологии и медицине в 1974 году за открытие рибосомы. Нобелевская премия по химии 2009 года присуждена Venkatraman Рамакришнан, Стейцам и Ад Е. Йонату для определения детальной структуры и механизма рибосомы.
 Состав рибосомной рРНК для прокариотической и эукариотической рРНК
Состав рибосомной рРНК для прокариотической и эукариотической рРНК 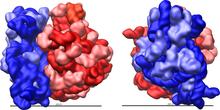 Рисунок 2: Большая (красная) и малая (синяя) субъединицы подходят друг к другу.
Рисунок 2: Большая (красная) и малая (синяя) субъединицы подходят друг к другу. Рибосома - сложная клеточная машина. Он в значительной степени состоит из специализированной РНК, известной как рибосомная РНК (рРНК), а также из десятков различных белков (точное количество незначительно варьируется между видами). Рибосомные белки и рРНК организованы в две отдельные рибосомные части разного размера, обычно известные как большая и малая субъединица рибосомы. Рибосомы состоят из двух субъединиц, которые подходят друг к другу (рис. 2) и работают как одна, транслируя мРНК в полипептидную цепь во время синтеза белка (рис. 1). Поскольку они образованы из двух субъединиц неравного размера, они немного длиннее по оси, чем по диаметру.
Бактериальные рибосомы имеют диаметр около 20 нм (200 Å ) и состоят из 65% рРНК и 35% рибосомных белков. Эукариотические рибосомы имеют диаметр от 25 до 30 нм (250–300 Å) с отношением рРНК к белку, близким к 1. Кристаллографические исследования показали, что поблизости от места реакции синтеза полипептидов нет рибосомных белков. Это говорит о том, что белковые компоненты рибосом не участвуют напрямую в катализе образования пептидных связей, а, скорее, эти белки действуют как каркас, который может повышать способность рРНК синтезировать белок (см. Рибозим ).
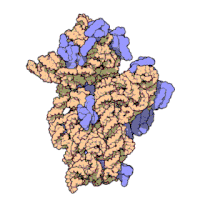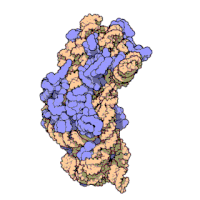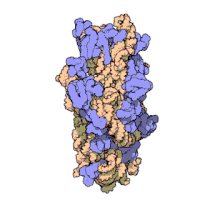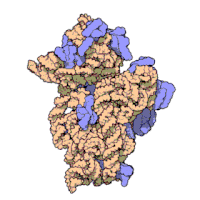 Рисунок 3: Атомная структура 30S субъединицы из Thermus thermophilus. Белки показаны синим цветом, а одиночная цепь РНК - коричневым.
Рисунок 3: Атомная структура 30S субъединицы из Thermus thermophilus. Белки показаны синим цветом, а одиночная цепь РНК - коричневым. Рибосомные субъединицы бактерий и эукариот очень похожи.
Единицей измерения, используемой для описания рибосомных субъединиц и фрагментов рРНК, является единица Сведберга, мера скорости оседания при центрифугировании, а не размера. Это объясняет, почему имена фрагментов не складываются: например, бактериальные 70S рибосомы состоят из 50S и 30S субъединиц.
Бактерии имеют 70 S рибосом, каждая из которых состоит из маленькой ( 30S ) и большой ( 50S ) субъединицы. E. coli, например, имеет субъединицу 16S РНК (состоящую из 1540 нуклеотидов), которая связана с 21 белком. Большая субъединица состоит из субъединицы 5S РНК (120 нуклеотидов), субъединицы 23S РНК (2900 нуклеотидов) и 31 белка.
| рибосома | подразделение | рРНК | р-белки |
|---|---|---|---|
| 70-е годы | 50S | 23S (2904 нт ) | 31 год |
| 5S (120 нт) | |||
| 30S | 16S (1542 нт) | 21 год |
Аффинная метка для сайтов связывания тРНК на рибосоме E. coli позволила идентифицировать белки сайтов A и P, наиболее вероятно связанные с пептидилтрансферазной активностью; меченые белки - L27, L14, L15, L16, L2; по крайней мере, L27 расположен на донорском участке, как показано E. Collatz и AP Czernilofsky. Дополнительные исследования показали, что белки S1 и S21 в ассоциации с 3'-концом 16S рибосомной РНК участвуют в инициации трансляции.
Рибосомы архей имеют те же общие размеры, что и бактериальные, и представляют собой рибосомы 70S, состоящие из большой субъединицы 50S, малой субъединицы 30S и содержащие три цепи рРНК. Однако на уровне последовательностей они намного ближе к эукариотическим, чем к бактериальным. Каждый дополнительный рибосомный белок архей по сравнению с бактериями имеет эукариотический аналог, в то время как между археями и бактериями такая связь отсутствует.
У эукариот в цитозоле расположены 80S рибосомы, каждая из которых состоит из маленькой (40S) и большой (60S) субъединицы. Их субъединица 40S содержит 18S РНК (1900 нуклеотидов) и 33 белка. Большая субъединица состоит из субъединиц 5S РНК (120 нуклеотидов), 28S РНК (4700 нуклеотидов), 5,8S РНК (160 нуклеотидов) и 46 белков.
| рибосома | подразделение | рРНК | р-белки |
|---|---|---|---|
| 80-е годы | 60S | 28S (4718 нт) | 49 |
| 5.8S (160 нт) | |||
| 5S (120 нт) | |||
| 40S | 18S (1874 г.) | 33 |
В 1977 году Черниловски опубликовал исследование, в котором использовалось аффинное мечение для определения сайтов связывания тРНК на рибосомах печени крысы. Некоторые белки, включая L32 / 33, L36, L21, L23, L28 / 29 и L13, были вовлечены как находящиеся в центре пептидилтрансферазы или рядом с ним.
У эукариот рибосомы присутствуют в митохондриях (иногда называемых миторибосомами ) и в пластидах, таких как хлоропласты (также называемые пласторибосомами). Они также состоят из больших и малых субъединиц, связанных вместе с белками в одну частицу 70S. Эти рибосомы похожи на рибосомы бактерий, и считается, что эти органеллы возникли как симбиотические бактерии. Из этих двух рибосомы хлоропластов ближе к бактериальным, чем митохрондриальные. Многие фрагменты рибосомальной РНК в митохрондриях укорачиваются, а в случае 5S рРНК у животных и грибов заменяются другими структурами. В частности, Leishmania tarentolae имеет минимизированный набор митохондриальной рРНК. Напротив, миторибосомы растений имеют как расширенную рРНК, так и дополнительные белки по сравнению с бактериями, в частности, многие повторяющиеся белки пентатрикопетида.
Криптофитовые водоросли и chlorarachniophyte водоросли могут содержать nucleomorph, который напоминает рудиментарное эукариотическое ядро. Рибосомы 80S эукариот могут присутствовать в компартменте, содержащем нуклеоморф.
Различия между бактериальными и эукариотическими рибосомами используются химиками-фармацевтами для создания антибиотиков, которые могут уничтожить бактериальную инфекцию, не нанося вреда клеткам инфицированного человека. Из-за различий в своей структуре бактериальные 70S рибосомы уязвимы для этих антибиотиков, в то время как эукариотические 80S рибосомы - нет. Несмотря на то, что митохондрии обладают рибосомами, подобными бактериальным, митохондрии не подвержены воздействию этих антибиотиков, потому что они окружены двойной мембраной, которая не позволяет этим антибиотикам легко проникать в органеллы. Однако заслуживающий внимания контрпример включает противоопухолевый антибиотик хлорамфеникол, который успешно ингибирует бактериальные 50S и митохондриальные 50S рибосомы эукариот. То же самое о митохондриях нельзя сказать о хлоропластах, где устойчивость рибосомных белков к антибиотикам является признаком, который следует ввести в качестве маркера в генную инженерию.
Различные рибосомы имеют общую структуру ядра, которая очень похожа, несмотря на большие различия в размерах. Большая часть РНК высокоорганизована в различные третичные структурные мотивы, например псевдоузлы, которые демонстрируют коаксиальный стэкинг. Дополнительная РНК в более крупных рибосомах находится в виде нескольких длинных непрерывных вставок, так что они образуют петли вне основной структуры, не нарушая и не изменяя ее. Вся каталитическая активность рибосомы осуществляется РНК ; белки находятся на поверхности и, кажется, стабилизируют структуру.
 Рисунок 4: Атомная структура субъединицы 50S из Haloarcula marismortui. Белки показаны синим, а две цепи РНК - коричневым и желтым. Небольшое зеленое пятно в центре субъединицы - активный сайт.
Рисунок 4: Атомная структура субъединицы 50S из Haloarcula marismortui. Белки показаны синим, а две цепи РНК - коричневым и желтым. Небольшое зеленое пятно в центре субъединицы - активный сайт. Общая молекулярная структура рибосомы известна с начала 1970-х годов. В начале 2000-х годов структура была достигнута при высоком разрешении порядка нескольких ангстрёмов.
Первые статьи, описывающие структуру рибосомы при атомном разрешении, были опубликованы почти одновременно в конце 2000 года. Субъединица 50S (большая прокариотическая) была определена из архей Haloarcula marismortui и бактерии Deinococcus radiodurans, а структура 30S-субъединицы была определена из Thermus thermophilus. Эти структурные исследования были удостоены Нобелевской премии по химии в 2009 году. В мае 2001 года эти координаты были использованы для реконструкции всей частицы T. thermophilus 70S с разрешением 5,5 Å.
В ноябре 2005 г. были опубликованы две статьи со структурами 70S рибосомы Escherichia coli. Структуры вакантной рибосомы определяли с разрешением 3,5 Å с помощью рентгеновской кристаллографии. Затем, две недели спустя, была опубликована структура, основанная на криоэлектронной микроскопии, которая изображает рибосому с разрешением 11-15 Å в процессе прохождения вновь синтезированной белковой цепи в белок-проводящий канал.
Первые атомные структуры рибосомы в комплексе с молекулами тРНК и мРНК были решены с помощью рентгеновской кристаллографии двумя группами независимо друг от друга: 2,8 Å и 3,7 Å. Эти структуры позволяют увидеть детали взаимодействия рибосомы Thermus thermophilus с мРНК и с тРНК, связанными в классических рибосомных сайтах. Взаимодействия рибосомы с длинными мРНК, содержащими последовательности Шайна-Дальгарно, были визуализированы вскоре после этого с разрешением 4,5–5,5 Å.
В 2011 году с помощью кристаллографии была получена первая полная атомная структура эукариотической рибосомы 80S дрожжей Saccharomyces cerevisiae. Модель раскрывает архитектуру эукариот-специфичных элементов и их взаимодействие с универсально сохраняемым ядром. В то же время была опубликована полная модель эукариотической рибосомной структуры 40S у Tetrahymena thermophila, описывающая структуру субъединицы 40S, а также многое о взаимодействии субъединицы 40S с eIF1 во время инициации трансляции. Аналогичным образом, структура эукариотической субъединицы 60S была также определена из Tetrahymena thermophila в комплексе с eIF6.
Рибосомы - это мельчайшие частицы, состоящие из РНК и связанных белков, которые функционируют для синтеза белков. Белки необходимы для многих клеточных функций, таких как восстановление повреждений или управление химическими процессами. Рибосомы можно обнаружить плавающими в цитоплазме или прикрепленными к эндоплазматической сети. Их основная функция - преобразование генетического кода в аминокислотную последовательность и построение белковых полимеров из аминокислотных мономеров.
Рибосомы действуют как катализаторы в двух чрезвычайно важных биологических процессах, называемых переносом пептидила и гидролизом пептидила. «Центр PT отвечает за образование белковых связей во время удлинения белка».
Рибосомы - это место биосинтеза белка, процесса трансляции мРНК в белок. МРНК содержит серию кодонов, которые декодируются рибосомой для образования белка. Используя мРНК в качестве матрицы, рибосома пересекает каждый кодон (3 нуклеотида ) мРНК, спаривая его с соответствующей аминокислотой, обеспечиваемой аминоацил-тРНК. Аминоацил-тРНК содержит комплементарный антикодон на одном конце и соответствующую аминокислоту на другом. Для быстрого и точного распознавания соответствующей тРНК рибосома использует большие конформационные изменения ( конформационная корректура ). Маленькая рибосомная субъединица, обычно связанная с аминоацил-тРНК, содержащей первую аминокислоту метионин, связывается с кодоном AUG на мРНК и рекрутирует большую рибосомную субъединицу. Рибосома содержит три сайта связывания РНК, обозначенных A, P и E. Сайт A связывает аминоацил-тРНК или факторы терминации высвобождения; Р-сайт связывает пептидил-тРНК (тРНК связывается с поли-пептидной цепи); и E-сайт (выход) связывает свободную тРНК. Синтез белка начинается со стартового кодона AUG около 5'-конца мРНК. мРНК сначала связывается с Р-сайтом рибосомы. Рибосома распознает стартовый кодон с помощью последовательности мРНК Шайна-Далгарно у прокариот и бокса Козака у эукариот.
Хотя в катализе пептидной связи участвует C2- гидроксил аденозина P-участка РНК в механизме протонного челнока, другие этапы синтеза белка (например, транслокация) вызываются изменениями в конформациях белка. Поскольку их каталитическое ядро состоит из РНК, рибосомы классифицируются как « рибозимы », и считается, что они могут быть остатками мира РНК.
 Рисунок 5: Трансляция мРНК (1) рибосомой (2) (показаны как малые и большие субъединицы) в полипептидную цепь (3). Рибосома начинается в стартовом кодоне РНК ( AUG) и заканчивается стоп-кодоном ( UAG).
Рисунок 5: Трансляция мРНК (1) рибосомой (2) (показаны как малые и большие субъединицы) в полипептидную цепь (3). Рибосома начинается в стартовом кодоне РНК ( AUG) и заканчивается стоп-кодоном ( UAG). На рисунке 5 обе субъединицы рибосомы ( малая и большая) собираются в стартовом кодоне (ближе к 5'-концу мРНК ). Рибосома использует тРНК, которая соответствует текущему кодону (триплету) на мРНК, чтобы присоединить аминокислоту к полипептидной цепи. Это делается для каждого триплета мРНК, в то время как рибосома перемещается к 3'-концу мРНК. Обычно в бактериальных клетках несколько рибосом работают параллельно с одной мРНК, образуя так называемую полирибосому или полисому.
Известно, что рибосома активно участвует в сворачивании белка. Полученные таким образом структуры обычно идентичны структурам, полученным в ходе химического рефолдинга белка; однако пути, ведущие к конечному продукту, могут быть разными. В некоторых случаях рибосома имеет решающее значение для получения функциональной белковой формы. Например, один из возможных механизмов сворачивания белков с глубокими узлами основан на том, что рибосома проталкивает цепь через прикрепленную петлю.
Присутствие белка контроля качества рибосом Rqc2 связано с мРНК-независимым удлинением белка. Это удлинение является результатом того рибосомального (через тРНК, принесенный Rqc2) из CAT хвостов: рибосомы расширить C -terminus из остановленного белка со случайными, переводом-независимых последовательностями через lanines и т hreonines.
Рибосомы классифицируются как «свободные» или «связанные с мембраной».
Свободные и связанные с мембраной рибосомы различаются только своим пространственным распределением; они идентичны по структуре. Независимо от того, существует ли рибосома в свободном или мембраносвязанном состоянии, зависит от присутствия на синтезируемом белке сигнальной последовательности, нацеленной на ER, поэтому отдельная рибосома может быть связана с мембраной, когда она производит один белок, но свободна в цитозоле. когда он производит другой белок.
Рибосомы иногда называют органеллами, но использование термина « органелла» часто ограничивается описанием субклеточных компонентов, которые включают фосфолипидную мембрану, а рибосомы, будучи полностью состоящими из частиц, не имеют. По этой причине рибосомы иногда называют «немембранными органеллами».
Свободные рибосомы могут перемещаться в любом месте цитозоля, но исключены из ядра клетки и других органелл. Белки, образованные из свободных рибосом, попадают в цитозоль и используются внутри клетки. Поскольку цитозоль содержит высокие концентрации глутатиона и, следовательно, является восстанавливающей средой, белки, содержащие дисульфидные связи, которые образуются из окисленных остатков цистеина, не могут продуцироваться в нем.
Когда рибосома начинает синтезировать белки, которые необходимы некоторым органеллам, рибосома, производящая этот белок, может стать «связанной с мембраной». В эукариотических клетках это происходит в области эндоплазматического ретикулума (ER), называемой «грубым ER». Вновь полученные полипептидные цепи вставляются непосредственно в ER рибосомой, осуществляющей векторный синтез, и затем транспортируются к месту назначения через секреторный путь. Связанные рибосомы обычно производят белки, которые используются внутри плазматической мембраны или выводятся из клетки посредством экзоцитоза.
В бактериальных клетках рибосомы синтезируются в цитоплазме посредством транскрипции оперонов множественных рибосомных генов. У эукариот этот процесс происходит как в цитоплазме клетки, так и в ядрышке, которое является областью внутри ядра клетки. Процесс сборки включает скоординированную функцию более 200 белков в синтезе и процессинге четырех рРНК, а также сборку этих рРНК с рибосомными белками.
Рибосома, возможно, сначала возникла в мире РНК, появившись как самовоспроизводящийся комплекс, который только позже развил способность синтезировать белки, когда начали появляться аминокислоты. Исследования показывают, что древние рибосомы, построенные исключительно из рРНК, могли развить способность синтезировать пептидные связи. Кроме того, данные убедительно указывают на древние рибосомы как самовоспроизводящиеся комплексы, в которых рРНК в рибосомах имела информационные, структурные и каталитические цели, поскольку она могла кодировать тРНК и белки, необходимые для саморепликации рибосом. Гипотетические клеточные организмы с самовоспроизводящейся РНК, но без ДНК называются рибоцитами (или рибоцеллами).
Поскольку аминокислоты постепенно появлялись в мире РНК в пребиотических условиях, их взаимодействие с каталитической РНК увеличивало как диапазон, так и эффективность функции молекул каталитической РНК. Таким образом, движущей силой эволюции рибосомы из древней самовоспроизводящейся машины в ее нынешнюю форму в качестве трансляционной машины могло быть давление отбора, направленное на включение белков в механизмы самовоспроизведения рибосомы, с тем чтобы увеличить ее способность к самовоспроизведение.
Рибосомы неоднородны по составу между видами и даже внутри одной клетки, о чем свидетельствует существование цитоплазматических и митохондриальных рибосом в одних и тех же эукариотических клетках. Некоторые исследователи предположили, что гетерогенность в составе рибосомных белков у млекопитающих важна для регуляции генов, т. Е. Гипотеза о специализированных рибосомах. Однако эта гипотеза является спорной и является предметом текущих исследований.
Неоднородность в составе рибосом была впервые предложена Винсом Мауро и Джеральдом Эдельманом для трансляционного контроля синтеза белка. Они предложили гипотезу рибосомного фильтра для объяснения регуляторных функций рибосом. Данные свидетельствуют о том, что специализированные рибосомы, специфичные для разных популяций клеток, могут влиять на то, как транслируются гены. Некоторые рибосомные белки обмениваются из собранного комплекса с цитозольными копиями, предполагая, что структура рибосомы in vivo может быть изменена без синтеза целой новой рибосомы.
Некоторые рибосомальные белки абсолютно необходимы для клеточной жизни, а другие нет. У почкующихся дрожжей 14/78 рибосомных белков не являются необходимыми для роста, тогда как у людей это зависит от исследуемой клетки. Другие формы гетерогенности включают посттрансляционные модификации рибосомных белков, такие как ацетилирование, метилирование и фосфорилирование. Arabidopsis, вирусные внутренние сайты входа в рибосомы (IRES) могут опосредовать трансляции различными по составу рибосомами. Например, 40S рибосомные единицы без eS25 в клетках дрожжей и млекопитающих неспособны рекрутировать CrPV IGR IRES.
Гетерогенность модификаций рибосомной РНК играет важную роль в поддержании структуры и / или функции, и большинство модификаций мРНК обнаруживаются в высококонсервативных областях. Наиболее распространенными модификациями рРНК являются псевдоуридилирование и 2'-O-метилирование рибозы.