
Войти
 Цитозоль - это переполненный раствор многих различных типов молекул, которые занимают до 30% цитоплазматической
Цитозоль - это переполненный раствор многих различных типов молекул, которые занимают до 30% цитоплазматической | Клеточная биология | |
|---|---|
| животная клетка | |
 Компоненты типичной животной клетки: Компоненты типичной животной клетки:
|
цитозоль, также известный как внутриклеточная жидкость (ICF ) или цитоплазматический матрикс, или основная плазма - это жидкость, находящаяся внутри клеток. Она разделена на отсеки мембраны. Например, митохондриальный матрикс разделяет митохондрию на множество компартментов. нц.
В эукариотической клетке цитозоль окружен клеточной мембраной и является частью цитоплазмы, которая также включает митохондрии, пластиды и другие органеллы (но не их внутренние жидкости и структуры); клеточное ядро является отдельным. Таким образом, цитозоль представляет собой жидкую матрицу вокруг органелл. У прокариот большинство химических реакций метаболизма происходит в цитозоле, а некоторые - в мембранах или в периплазматическом пространстве. У эукариот, хотя многие метаболические пути все еще происходят в цитозоле, другие происходят внутри органелл.
Цитозоль представляет собой сложную смесь веществ, растворенных в воде. Хотя вода составляет большую часть цитозоля, ее структура и свойства внутри клеток изучены недостаточно. Концентрации ионов, таких как натрия и калий, отличаются в цитозоле от внеклеточной жидкости ; эти различия в уровнях ионов важны для таких процессов, как осморегуляция, передача сигналов клеток и генерация потенциалов действия в возбудимых клетках, таких как эндокринные, нервные и мышечные клетки. Цитозоль также содержит большое количество макромолекул, которые могут изменять поведение молекул за счет макромолекулярного скопления.
Хотя когда-то считалось, что это простой раствор молекул, цитозоль имеет несколько уровней организация. К ним относятся градиенты концентрации малых молекул, таких как кальций, большие комплексы ферментов, которые действуют вместе и участвуют в метаболических путях, и белковые комплексы, такие как протеасомы и карбоксисомы, которые охватывают и разделяют части цитозоля.
Впервые был введен термин «цитозоль» в 1965 г. HA Lardy и первоначально относился к жидкости, которая была получена путем разрушения клеток и осаждения всех нерастворимых компонентов с помощью ультрацентрифугирования. Такой растворимый клеточный экстракт не идентичен растворимой части цитоплазмы клетки и обычно называется цитоплазматической фракцией.
Термин цитозоль теперь используется для обозначения жидкой фазы цитоплазмы в интактной клетке. Это исключает любую часть цитоплазмы, которая содержится в органеллах. Из-за возможности путаницы между использованием слова «цитозоль» для обозначения как экстрактов клеток, так и растворимой части цитоплазмы в интактных клетках фраза «водная цитоплазма» использовалась для описания жидкого содержимого цитоплазмы. живых клеток.
До этого другие термины, включая гиалоплазма, использовались для обозначения клеточной жидкости, не всегда как синонимы, поскольку ее природа была не очень ясна (см. протоплазма ).
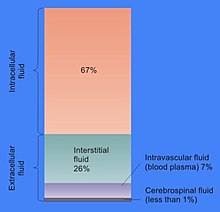 Содержание внутриклеточной жидкости у человека
Содержание внутриклеточной жидкости у человека Пропорция объема клетки, которая составляет цитозоль, варьируется: например, в то время как этот компартмент формирует основную часть клеточной структуры в бактериях, в клетках растений основной отсек - большая центральная вакуоль. Цитозоль состоит в основном из воды, растворенных ионов, небольших молекул и крупных водорастворимых молекул (например, белков). Большинство этих небелковых молекул имеют молекулярная масса менее 300 Да. Эта смесь небольших молекулы чрезвычайно сложны, так как разнообразие молекул, участвующих в метаболизме (метаболиты ), огромно. Например, в растениях может образовываться до 200 000 различных небольших молекул, хотя не все они будут присутствовать в одном и том же виде или в одной клетке. Оценки количества метаболитов в отдельных клетках, таких как E. coli и хлебопекарные дрожжи предсказывают, что производится менее 1000.
Большая часть цитозоля - это вода, что составляет около 70% от общего объема типовой ячейки. pH внутриклеточной жидкости составляет 7,4. в то время как цитозольный pH человека находится в диапазоне 7,0–7,4 и обычно выше, если клетка растет. вязкость цитоплазмы примерно такая же, как у чистой воды, хотя диффузия небольших молекул через эту жидкость примерно в четыре раза медленнее, чем в чистой воде, в основном из-за столкновений с большим количеством макромолекулы в цитозоле. Исследования на морских креветках изучали, как вода влияет на функции клеток; они увидели, что уменьшение количества воды в клетке на 20% подавляет метаболизм, причем метаболизм постепенно снижается по мере высыхания клетки, и вся метаболическая активность останавливается, когда уровень воды достигает 70% ниже нормы.
Хотя вода жизненно важна для жизни, структура этой воды в цитозоле недостаточно изучена, в основном потому, что такие методы, как спектроскопия ядерного магнитного резонанса, дают информацию только о средней структуре воды и не могут измерять локальные изменения в микроскопическая шкала. Даже структура чистой воды плохо изучена из-за способности воды образовывать структуры, такие как кластеры воды через водородные связи.
Классический взгляд на воду в клетках заключается в том, что около 5% Эта вода прочно связана растворенными веществами или макромолекулами, как вода сольватации, в то время как большая часть имеет такую же структуру, как чистая вода. Эта сольватационная вода не активна в осмосе и может иметь другие свойства растворителя, так что некоторые растворенные молекулы исключаются, а другие становятся концентрированными. Однако другие утверждают, что влияние высоких концентраций макромолекул в клетках распространяется на весь цитозоль и что вода в клетках ведет себя совершенно иначе, чем вода в разбавленных растворах. Эти идеи включают предположение, что клетки содержат зоны с низкой и высокой плотностью воды, которые могут иметь широкое влияние на структуры и функции других частей клетки. Однако использование передовых методов ядерного магнитного резонанса для прямого измерения подвижности воды в живых клетках противоречит этой идее, поскольку предполагает, что 85% клеточной воды действует как чистая вода, а остальная часть менее подвижна и, вероятно, связана с макромолекулами..
Концентрации других ионов в цитозоле сильно отличаются от таковых во внеклеточной жидкости, и цитозоль также содержит гораздо большее количество заряженных макромолекул, таких как белки и нуклеиновые кислоты, чем вне клеточной структуры.
| Ионы | Концентрация (миллимолярная) | |
|---|---|---|
| В цитозоле | В плазме | |
| Калий | 139–150 | 4 |
| Натрий | 12 | 145 |
| Хлорид | 4 | 116 |
| Бикарбонат | 12 | 29 |
| Аминокислоты в белках | 138 | 9 |
| Магний | 0,8 | 1,5 |
| Кальций | <0.0002 | 1,8 |
В отличие от внеклеточной жидкости, цитозоль имеет высокую концентрацию ионов калия и низкая концентрация ионов натрия. Эта разница в концентрациях ионов имеет решающее значение для осморегуляции, поскольку, если бы уровни ионов внутри клетки были такими же, как и снаружи, вода поступала бы постоянно из-за осмоса - поскольку уровни макромолекулы внутри клеток выше, чем их уровни снаружи. Вместо этого ионы натрия вытесняются, а ионы калия поглощаются Na⁺ / K⁺-АТФазой, ионы калия затем стекают вниз по градиенту их концентрации через ионные каналы отбора калия, эта потеря положительного заряда создает отрицательный мембранный потенциал. Чтобы уравновесить эту разность потенциалов, отрицательные ионы хлора также выходят из ячейки через селективные хлоридные каналы. Потеря ионов натрия и хлорида компенсирует осмотический эффект более высокой концентрации органических молекул внутри клетки.
Клетки могут справляться даже с более значительными осмотическими изменениями, накапливая осмозащитные вещества, такие как бетаины или трегалоза в их цитозоле. Некоторые из этих молекул могут позволить клеткам выжить, будучи полностью высохшими, и позволяют организму войти в состояние анабиоза, называемое криптобиозом. В этом состоянии цитозоль и осмопротекторы превращаются в стеклообразное твердое вещество, которое помогает стабилизировать белки и клеточные мембраны от разрушающего воздействия высыхания.
Низкая концентрация кальция в цитозоле позволяет ионам кальция функционировать как второй посланник в кальциевой сигнализации. Здесь сигнал, такой как гормон или потенциал действия, открывает кальциевый канал, так что кальций наводняется в цитозоль. Это внезапное увеличение цитозольного кальция активирует другие сигнальные молекулы, такие как кальмодулин и протеинкиназа C. Другие ионы, такие как хлорид и калий, также могут выполнять сигнальные функции в цитозоле, но они недостаточно изучены.
Молекулы белка, которые не связываются с клеточными мембранами или цитоскелет растворяются в цитозоле. Количество белка в клетках чрезвычайно велико и приближается к 200 мг / мл, занимая около 20–30% объема цитозоля. Однако точно измерить, сколько белка растворено в цитозоле в интактных клетках, сложно, поскольку некоторые белки, по-видимому, слабо связаны с мембранами или органеллами в целых клетках и высвобождаются в раствор при лизисе клеток. Действительно, в экспериментах, где плазматическая мембрана клеток была осторожно разрушена с помощью сапонина, без повреждения других клеточных мембран, высвобождалась только около четверти клеточного белка. Эти клетки также были способны синтезировать белки, если им давали АТФ и аминокислоты, что означает, что многие ферменты в цитозоле связаны с цитоскелетом. Однако идея о том, что большинство белков в клетках прочно связаны в сеть, называемую микротрабекулярной решеткой, сейчас считается маловероятной.
У прокариот цитозоль содержит клеточные геном в структуре, известной как нуклеоид. Это нерегулярная масса ДНК и связанных белков, которые контролируют транскрипцию и репликацию бактериальной хромосомы и плазмид. У эукариот геном удерживается внутри клеточного ядра, которое отделено от цитозоля ядерными порами, которые блокируют свободную диффузию любой молекулы размером более примерно 10 нанометров в диаметре.
Эта высокая концентрация макромолекул в цитозоле вызывает эффект, называемый макромолекулярным скоплением, когда эффективная концентрация других макромолекул увеличивается, поскольку они имеют меньший объем для проникновения. Этот эффект вытеснения может вызвать большие изменения как в скоростях, так и в положении химического равновесия реакций в цитозоле. Это особенно важно из-за его способности изменять константы диссоциации, способствуя ассоциации макромолекул, например, когда несколько белков объединяются, образуя белковые комплексы, или когда связывание с ДНК белки связываются со своими мишенями в геноме.
Хотя компоненты цитозоля не разделены на участки клеточными мембранами, эти компоненты не всегда смешиваются случайным образом и несколько уровней Организация может локализовать определенные молекулы в определенных местах внутри цитозоля.
Хотя небольшие молекулы быстро диффундируют в цитозоле, градиенты концентрации все же могут возникать в этом отсеке. Хорошо изученным примером этого являются «кальциевые искры», которые образуются в течение короткого периода в области вокруг открытого кальциевого канала. Они составляют около 2 микрометров в диаметре и длятся всего несколько миллисекунд, хотя несколько искр могут сливаться, образуя более крупные градиенты, называемые «кальциевыми волнами». Градиенты концентрации других малых молекул, таких как кислород и аденозинтрифосфат, могут образовываться в клетках вокруг кластеров митохондрий, хотя они менее изучены.
Белки могут связываться с образованием белковых комплексов, они часто содержат набор белков со схожими функциями, например ферменты, которые выполняют несколько этапов одного и того же метаболического пути.. Эта организация может позволить субстратный канал, когда продукт одного фермента передается непосредственно следующему ферменту в пути, не попадая в раствор. Каналирование может сделать путь более быстрым и эффективным, чем если бы ферменты были беспорядочно распределены в цитозоле, а также может предотвратить высвобождение нестабильных промежуточных продуктов реакции. Хотя большое количество метаболических путей включает ферменты, которые тесно связаны друг с другом, другие могут включать более слабо связанные комплексы, которые очень трудно изучать вне клетки. Следовательно, важность этих комплексов для метаболизма в целом остается неясной.
 Карбоксисомы представляют собой заключенные в белок бактериальные микрокомпартменты внутри цитозоля. Слева электронное микроскопическое изображение карбоксисом, а справа - модель их структуры.
Карбоксисомы представляют собой заключенные в белок бактериальные микрокомпартменты внутри цитозоля. Слева электронное микроскопическое изображение карбоксисом, а справа - модель их структуры. Некоторые белковые комплексы содержат большую центральную полость, изолированную от остаток цитозоля. Одним из примеров такого закрытого отсека является протеасома. Здесь набор субъединиц образует полый цилиндр, содержащий протеазы, которые разрушают цитозольные белки. Поскольку они могли бы повредить, если бы они свободно смешивались с остальной частью цитозоля, ствол закрыт набором регуляторных белков, которые распознают белки с сигналом, направляющим их на деградацию (тег убиквитин ), и питают их. в протеолитическую полость.
Другой большой класс белковых компартментов - это бактериальные микрокомпартменты, которые состоят из белковой оболочки, которая инкапсулирует различные ферменты. Эти компартменты обычно имеют размер около 100–200 нанометров в поперечнике и состоят из взаимосвязанных белков. Хорошо понятным примером является карбоксисома, которая содержит ферменты, участвующие в фиксации углерода, такие как RuBisCO.
немембранные органеллы. могут образовываться в виде биомолекулярных конденсатов, которые возникают в результате кластеризации, олигомеризации или полимеризации макромолекул с образованием коллоидных фазовое разделение цитоплазмы или ядра.
Хотя цитоскелет не является частью цитозоля, присутствие этой сети нитей ограничивает диффузию крупных частиц в клетке. Например, в нескольких исследованиях индикаторные частицы размером более 25 нанометров (размером с рибосому ) были исключены из частей цитозоля по краям клетки и рядом с ней. ядро. Эти «исключающие компартменты» могут содержать гораздо более плотную сетку из волокон актина, чем остальная часть цитозоля. Эти микродомены могут влиять на распределение крупных структур, таких как рибосомы и органеллы в цитозоле, исключая их из одних областей и концентрируя их в других.
Цитозоль не имеет единой функции и вместо этого является участком множества клеточных процессов. Примеры этих процессов включают передачу сигнала от клеточной мембраны к участкам внутри клетки, таким как ядро клетки или органеллы. Этот компартмент также является местом многих процессов цитокинеза после разрушения ядерной мембраны в митозе. Другой важной функцией цитозоля является транспортировка метаболитов от места их производства к месту их использования. Это относительно просто для водорастворимых молекул, таких как аминокислоты, которые могут быстро диффундировать через цитозоль. Однако гидрофобные молекулы, такие как жирные кислоты или стерины, могут транспортироваться через цитозоль специфическими связывающими белками, которые перемещают эти молекулы между клеточными мембранами. Молекулы, попавшие в клетку в результате эндоцитоза или находящиеся на пути секретирования, также могут транспортироваться через цитозоль внутри везикул, которые представляют собой небольшие сферы липидов, которые перемещается вдоль цитоскелета с помощью моторных белков.
Цитозоль является местом наибольшего метаболизма у прокариот и значительной части метаболизма эукариот. Например, у млекопитающих около половины белков в клетке локализованы в цитозоле. Наиболее полные данные доступны по дрожжам, где метаболические реконструкции показывают, что большинство метаболических процессов и метаболитов происходит в цитозоле. Основные метаболические пути, которые происходят в цитозоле у животных, - это биосинтез белка, пентозофосфатный путь, гликолиз и глюконеогенез. Локализация путей может отличаться у других организмов, например, синтез жирных кислот происходит в хлоропластах у растений и в апикопластах в apicomplexa.