
Войти
Секция - это перемещение материала из одной точки в другую, например, секретируемого химического вещества из ячейка или сальник. Напротив, экскреция - это удаление определенных веществ или продуктов жизнедеятельности из клетки или организма. Классический механизм секреции клеток осуществляется через секреторные порталы на плазматической мембране клетки, которые называются поросомами. Поросомы представляют собой постоянную чашеобразную липопротеиновую структуру на плазматической мембране клетки, где секреторные везикулы временно стыкуются и сливаются, высвобождая внутривезикулярное содержимое из клетки.
Секреция у видов бактерий означает транспорт или транслокацию эффекторных молекул, например: белков, ферментов или токсинов (таких как токсин холеры в патогенных бактериях, например Vibrio cholerae ) изнутри (цитоплазма или цитозоль ) бактериальной клетки к его внешности. Секреция - очень важный механизм функционирования бактерий и их функционирования в их естественной окружающей среде для адаптации и выживания.
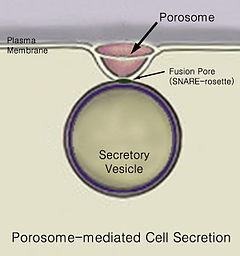 Поросома
Поросома Эукариотические клетки, включая клетки человека, имеют высоко развитый процесс секреции. Белки , нацеленные извне, синтезируются с помощью рибосом, прикрепленных к грубому эндоплазматическому ретикулуму (ER). По мере синтеза эти белки перемещаются в просвет ER, где они гликозилированы и где молекулярные шапероны помогают сворачиванию белка. Неправильно свернутые белки здесь обычно идентифицируются и ретранслируются посредством ER-ассоциированной деградации в цитозоль, где они расщепляются протеасомой. везикулы, содержащие правильно уложенные белки, затем попадают в аппарат Гольджи.
. В аппарате Гольджи гликозилирование белков модифицируется и происходит дальнейшие посттрансляционные модификации, включая расщепление и функционализация, может произойти. Затем белки перемещаются в секреторные пузырьки, которые перемещаются вдоль цитоскелета к краю клетки. Дополнительная модификация может происходить в секреторных везикулах (например, инсулин отщепляется от проинсулина в секреторных везикулах).
В конце концов, происходит слияние пузырьков с клеточной мембраной в структуре, называемой поросомой, в процессе, называемом экзоцитоз, выбрасывая его содержимое из среды клетки.
Строгий биохимический контроль над этой последовательностью поддерживается за счет использования градиента pH : pH цитозоля составляет 7,4, pH ER составляет 7,0, а pH цис-гольджи составляет 6,5. Секреторные везикулы имеют pH от 5,0 до 6,0; некоторые секреторные везикулы превращаются в лизосомы с pH 4,8.
Существует множество белков, таких как FGF1 (aFGF), FGF2 (bFGF), интерлейкин-1 (IL1) и т.д., которые не имеют сигнальной последовательности. Они не используют классический путь ER-Golgi. Они секретируются различными неклассическими путями.
Описано по меньшей мере четыре неклассических (нетрадиционных) пути секреции белка. Они включают в себя 1) прямую транслокацию белков через плазматическую мембрану, вероятно, через мембранные транспортеры, 2) пузырек, 3) лизосомную секрецию и 4) высвобождение через экзосомы, происходящие из мультивезикулярных телец. Кроме того, белки могут высвобождаться из клеток в результате механических или физиологических повреждений и через нелетальные временные онкотические поры в плазматической мембране, вызванные промывкой клеток бессывороточной средой или буферами.
Многие типы клеток человека обладают способностью быть секреторными клетками. У них есть хорошо развитая эндоплазматическая сеть и аппарат Гольджи для выполнения этой функции. Ткани, которые производят секрецию, включают желудочно-кишечный тракт, который секретирует пищеварительные ферменты и желудочную кислоту, легкие, которые выделяют поверхностно-активные вещества и сальные железы, которые выделяют кожный жир для смазывания кожи и волос. Мейбомиевые железы в веке секретируют meibum для смазки и защиты глаза.
Секреция характерна не только для эукариот - она также присутствует у бактерий и архей. Переносчики типа кассеты связывания АТФ (ABC) являются общими для трех областей жизни. Некоторые секретируемые белки перемещаются через цитоплазматическую мембрану с помощью транслокона Sec, одной из двух систем транслокации, которая требует присутствия N-концевого сигнального пептида на секретируемом белке. Другие перемещаются через цитоплазматическую мембрану по пути двойной транслокации аргинина (Tat). Грамотрицательные бактерии имеют две мембраны, что топологически усложняет секрецию. У грамотрицательных бактерий существует как минимум шесть специализированных систем секреции. Многие секретируемые белки особенно важны в бактериальном патогенезе.
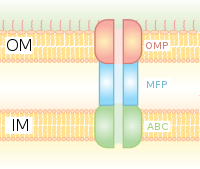
Секреция типа I представляет собой систему секреции, зависимую от шаперона, использующую кластеры генов Hly и Tol. Процесс начинается, когда лидерная последовательность HlyA распознается и связывает HlyB на мембране. Эта сигнальная последовательность чрезвычайно специфична для транспортера ABC. Комплекс HlyAB стимулирует HlyD, который начинает раскручиваться и достигает внешней мембраны, где TolC распознает концевую молекулу или сигнал на HlyD. HlyD рекрутирует TolC на внутреннюю мембрану, а HlyA выводится за пределы внешней мембраны через длинный туннельный белковый канал.
Система секреции типа I транспортирует различные молекулы, от ионов и лекарств до белков различного размера (20-900 кДа). Секретируемые молекулы различаются по размеру от небольшого пептида колицина V Escherichia coli (10 кДа) до белка клеточной адгезии LapA Pseudomonas fluorescens 520 кДа. Лучше всего охарактеризованы токсины RTX и липазы. Секреция типа I также участвует в экспорте небелковых субстратов, таких как циклические β-глюканы и полисахариды.

Белки, секретируемые через систему типа II или главную концевую ветвь общего секреторного пути, зависят от системы Sec или Tat для первоначального транспорта в периплазму. Оказавшись там, они проходят через внешнюю мембрану через мультимерный (12–14 субъединиц) комплекс порообразующих секретиновых белков. В дополнение к белку секретина, 10-15 других белков внутренней и внешней мембран составляют полный аппарат секреции, многие функции которых пока неизвестны. Грамотрицательные пили типа IV используют модифицированную версию системы типа II для своего биогенеза, и в некоторых случаях определенные белки являются общими между комплексом пилей и системой типа II в пределах одного вида бактерий.
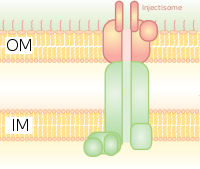
Она гомологична базальному тельцу бактериальных жгутиков. Это похоже на молекулярный шприц, через который бактерия (например, определенные типы Salmonella, Shigella, Yersinia, Vibrio ) могут вводить белки. в эукариотические клетки. Низкая концентрация Са в цитозоле открывает ворота, регулирующие T3SS. Один из таких механизмов обнаружения низкой концентрации кальция был проиллюстрирован антигеном lcrV (Low Calcium Response), используемым Yersinia pestis, который используется для обнаружения низких концентраций кальция и вызывает присоединение T3SS. Система Hrp в патогенах растений вводит в растения гарпины и эффекторные белки патогенов через аналогичные механизмы. Эта система секреции была впервые обнаружена у Yersinia pestis и показала, что токсины могут вводиться непосредственно из бактериальной цитоплазмы в цитоплазму клеток-хозяев, а не просто секретироваться во внеклеточную среду.
| T4SS | |||||||||
|---|---|---|---|---|---|---|---|---|---|
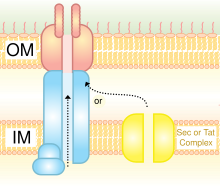 Система секреции типа IV Система секреции типа IV | |||||||||
| Идентификаторы | |||||||||
| Символ | T4SS | ||||||||
| Pfam | PF07996 | ||||||||
| InterPro | IPR012991 | ||||||||
| SCOPe | 1gl7 / SUPFAM | ||||||||
| TCDB | 3.A.7 | ||||||||
| суперсемейство OPM | 215 | ||||||||
| белок OPM | 3jqo | ||||||||
| |||||||||
Он гомологичен конъюгированному механизму бактерий. Он способен переносить как ДНК, так и белки. Он был обнаружен у Agrobacterium tumefaciens, который использует эту систему для введения части Т-ДНК плазмиды Ti в растение-хозяин, что, в свою очередь, приводит к тому, что пораженный участок превращается в коронный галл (опухоль). Helicobacter pylori использует систему секреции типа IV для доставки CagA в эпителиальные клетки желудка, что связано с канцерогенезом желудка. Bordetella pertussis, возбудитель коклюша, частично секретирует коклюшный токсин через систему IV типа. Legionella pneumophila, возбудитель легионеллеза (болезни легионеров), использует a, известный как icm / dot (i ntra c ellular m ультипликация / d эффект в системе o rganelle t rafficking genes) для перемещения множества эффекторных белков в его эукариотического хозяина. Прототипом системы секреции типа IVA является комплекс VirB Agrobacterium tumefaciens.
. Белки этого семейства являются компонентами системы секреции типа IV. Они опосредуют внутриклеточный перенос макромолекул через механизм , предков связанный с механизмом бактериальной конъюгации.
Короче говоря, система секреции типа IV (T4SS) - это общий механизм, с помощью которого бактериальные клетки секретируют или поглощают макромолекулы. Их точный механизм остается неизвестным. T4SS кодируется на грамотрицательных конъюгативных элементах в бактериях. T4SS - это комплексы, охватывающие клеточную оболочку, или, другими словами, 11-13 ядерных белков, которые образуют канал, через который ДНК и белки могут путешествовать из цитоплазмы клетки-донора в цитоплазму клетки-реципиента. Кроме того, T4SS также секретирует белки факторов вирулентности непосредственно в клетки-хозяева, а также захватывает ДНК из среды во время естественной трансформации, что демонстрирует универсальность этого аппарата секреции макромолекул.
Как показано на приведенном выше рисунке, TraC, в частности, состоит из трехспирального пучка и рыхлого шаровидного отростка.
T4SS имеет два эффектора белки: во-первых, ATS-1, который означает транслоцированный субстрат 1 из анаплазмы, и, во-вторых, AnkA, который означает белок A, содержащий домен анкиринового повтора. Кроме того, связывающими белками T4SS являются VirD4, которые связываются с VirE2.
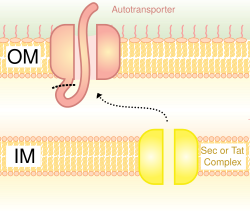
Также называемая системой аутотранспортера, секреция типа V включает использование системы Sec для пересечения внутренней мембраны. Белки, использующие этот путь, обладают способностью образовывать бета-ствол со своим С-концом, который вставляется во внешнюю мембрану, позволяя остальной части пептида (домен-пассажира) достигать внешней части клетки. Часто аутотранспортеры расщепляются, оставляя бета-бочкообразный домен на внешней мембране и освобождая пассажирский домен. Некоторые исследователи полагают, что остатки автотранспортеров дали начало поринам, которые образуют похожие бета-цилиндрические структуры. Распространенным примером аутотранспортера, который использует эту систему секреции, является тримерный аутотранспортер Adhesins.
Системы секреции типа VI были первоначально идентифицированы в 2006 году группой из Джон Мекаланос из Гарвардской медицинской школы (Бостон, США) по двум бактериальным патогенам: Vibrio cholerae и Pseudomonas aeruginosa. Они были идентифицированы, когда мутации в генах Hcp и VrgG в Vibrio Cholerae привели к снижению вирулентности и патогенности. С тех пор системы секреции типа VI были обнаружены в четверти всех геномов протеобактерий, включая патогены животных, растений, человека, а также почвенные, экологические или морские бактерии. В то время как большинство ранних исследований секреции типа VI было сосредоточено на ее роли в патогенезе высших организмов, более поздние исследования предполагали более широкую физиологическую роль в защите от простых эукариотических хищников и ее роль во взаимодействиях между бактериями. Кластеры генов системы секреции типа VI содержат от 15 до более чем 20 генов, два из которых, Hcp и VgrG, как было показано, являются почти универсально секретируемыми субстратами системы. Структурный анализ этих и других белков в этой системе поразительно похож на хвостовой шип фага Т4, и считается, что активность системы функционально напоминает фаговую инфекцию.
Помимо использования перечисленных выше мультибелковых комплексов, грамотрицательные бактерии обладают другим методом высвобождения материала: образованием везикул наружной мембраны бактерий. Части внешней мембраны отщипываются, образуя наноразмерные сферические структуры, состоящие из богатого липополисахаридами липидного бислоя, охватывающего периплазматические материалы, и используются для перемещения мембранных пузырьков для манипулирования окружающей средой или вторжения в хозяина патоген интерфейс. Было обнаружено, что везикулы ряда видов бактерий содержат факторы вирулентности, некоторые обладают иммуномодулирующим действием, а некоторые могут непосредственно прилипать к клеткам-хозяевам и отравлять их. высвобождение везикул было продемонстрировано как общий ответ на стрессовые условия, процесс загрузки грузовых белков кажется избирательным.
У некоторых видов Staphylococcus и Streptococcus, вспомогательная секреторная система отвечает за экспорт гликопротеинов адгезии с высокой повторяемостью.
| Найдите раздел в Wiktionary, бесплатном словаре. |