
Войти
Системы секреции бактерий представляют собой белковые комплексы, присутствующие на клеточные мембраны бактерий для секреции веществ. В частности, они представляют собой клеточные устройства, используемые патогенными бактериями для выделения факторов вирулентности (в основном белков) для вторжения в клетки-хозяева. Их можно разделить на разные типы в зависимости от их конкретной структуры, состава и активности. Эти основные различия можно различить между грамотрицательными и грамположительными бактериями. Но классификация отнюдь не ясна и полна. Существует как минимум восемь типов, специфичных для грамотрицательных бактерий, четыре - для грамположительных бактерий и два общих для обоих. Как правило, белки могут секретироваться двумя разными способами. Один процесс представляет собой одностадийный механизм, в котором белки из цитоплазмы бактерий транспортируются и доставляются непосредственно через клеточную мембрану в клетку-хозяин. Другой включает двухэтапную активность, при которой белки сначала транспортируются из внутренней клеточной мембраны, затем откладываются в периплазме и, наконец, через внешнюю клеточную мембрану в клетку-хозяин.
 Комплемент. секретома, участвующего в процессе колонизации у монодермальных бактерий.
Комплемент. секретома, участвующего в процессе колонизации у монодермальных бактерий. 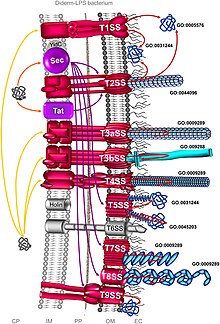 Системы секреции у дидермальных бактерий
Системы секреции у дидермальных бактерий Общая секреция (Sec) включает секрецию развернутых белков, которые сначала остаются внутри клеток. У грамотрицательных бактерий секретируемый белок направляется либо во внутреннюю мембрану, либо в периплазму. Но у грамположительных бактерий белок может оставаться в клетке или в основном выводится из бактерий с помощью других систем секреции. Среди грамотрицательных бактерий Vibrio cholerae, Klebsiella pneumoniae и Yersinia enterocolitica используют систему Sec. Staphylococcus aureus и Listeria monocytogenes - это грамположительные бактерии, которые используют систему Sec.
Система Sec использует два разных пути секреции: пути SecA и сигнальные частицы (SRP). SecA является моторным белком АТФазы и имеет множество родственных белков, включая SecD, SecE, SecF, SegG, SecM и SecY. SRP - это рибонуклеопротеин (комплекс белок-РНК), который распознает и нацеливает специфические белки на эндоплазматический ретикулум у эукариот и на клеточную мембрану у прокариот. Эти два пути требуют разных молекулярных шаперонов и, в конечном итоге, используют канал транспортировки белков SecYEG для транспортировки белков через внутреннюю клеточную мембрану. В пути SecA SecB действует как шаперон, помогая транспорту белка в периплазму после полного синтеза пептидных цепей. В то время как в пути SRP YidC является шапероном и транспортирует белки к клеточной мембране, пока они все еще подвергаются синтезу пептидов.
Белки синтезируются в рибосомах посредством процесс последовательного добавления аминокислот, называемый трансляцией. В пути SecA фактор запуска шаперона (TF) сначала связывается с экспонированной N-концевой сигнальной последовательностью пептидной цепи. По мере продолжения удлинения пептидной цепи ТФ заменяется на SecB. SecB специфически поддерживает пептид в развернутом состоянии и способствует связыванию SecA. Затем комплекс может связываться с SecYEG, посредством чего SecA активируется путем связывания с АТФ. Под действием энергии АТФ SecA проталкивает белок через канал secYEG. Комплекс SecD / F также помогает извлекать белок с другой стороны клеточной мембраны.
В этом пути SRP конкурирует с TF и связывается с N-концом сигнальная последовательность. Белки внутренней мембраны останавливают процесс удлинения цепи. Затем SRP связывается с мембранным рецептором FtsY. Комплекс пептидная цепь-SRP-FtsY затем транспортируется в SecY, где возобновляется удлинение пептида.
Система двойной транслокации аргинина (Tat) аналогична Sec в процессе образования белка. секреция, однако, он отправляет белки только в их свернутом (третичном) состоянии. Его используют все виды бактерий, а также археи, хлоропласты и митохондрии растений. У бактерий система Tat экспортирует белки из цитоплазмы через внутреннюю клеточную мембрану; тогда как в хлоропластах он присутствует в тилакоидной мембране, где он помогает импорту белков из стромы. Белки Tat очень вариабельны у разных бактерий и подразделяются на три основных типа, а именно TatA, TatB и TatC. Например, хотя в Bacillus subtilis есть только два функциональных белка Tat, у Streptomyces coelicolor их может быть более сотни. Сигнальные пептиды, которые могут распознавать белки Tat, характеризуются консенсусным мотивом Ser / Thr-Arg-Arg-X-Phe-Leu-Lys (где X может быть любой полярной аминокислотой). Это два последовательных аргинина, от которых произошло название «двойная транслокация аргинина». Замена любого из аргинина приводит к замедлению или нарушению секреции.
Система секреции типа I (T1SS или TOSS) обнаруживается у грамотрицательных бактерий. Это зависит от активности шаперона с использованием белков Hly и Tol. Система активируется, когда сигнальная последовательность HlyA связывает HlyB на клеточной мембране. Эта сигнальная последовательность является транспортером ABC. Комплекс HlyAB активирует HlyD, который разматывается и перемещается к внешней клеточной мембране. Конечный сигнал распознается TolC во внутренней мембране. HlyA секретируется из внешней мембраны через туннельный канал белка.
T1SS транспортирует различные молекулы, включая ионы, углеводы, лекарства, белки. Секретируемые молекулы различаются по размеру от небольшого пептида колицина V Escherichia coli, который составляет 10 кДа, до белка адгезии клеток Pseudomonas fluorescens LapA, который составляет 520 кДа. К наиболее известным молекулам относятся токсины RTX и ферменты липазы.
Система секреции типа II (T2SS) зависит от системы Sec или Tat для начальной секреции внутри бактериальной клетки. Из периплазмы белки секретируются из секретинов внешней мембраны. Секретины представляют собой мультимерный (12–14 субъединиц) комплекс порообразующих белков. Секретин поддерживается 10-15 другими белками внутренней и внешней мембраны, составляя полный аппарат секреции.
Система секреции типа III (T3SS или TTSS) структурно подобна и связана с базальное тело бактериальных жгутиков. Обнаруженный у некоторых из наиболее вирулентных грамотрицательных бактерий, таких как Salmonella, Shigella, Yersinia, Vibrio, он используется для инъекций токсичные белки в эукариотические клетки. По своей структуре его часто описывают как инъекционные или иглы и шприцы. Обнаружено с помощью Yersinia pestis, было обнаружено, что T3SS может вводить токсины непосредственно из бактериальной цитоплазмы в цитоплазму клеток своего хозяина.
Система секреции IV типа (T4SS или TFSS) относится к системе бактериальной конъюгации, с помощью которой различные бактерии могут обмениваться своими ДНК. Участвующие бактерии могут относиться к одному или к разным грамотрицательным видам бактерий. Он может транспортировать отдельные белки, а также комплексы белок-белок и ДНК-белок. Секреция передается непосредственно от клетки-реципиента через клеточные мембраны. Agrobacterium tumefaciens, из которой она была первоначально обнаружена, использует эту систему для отправки части Т-ДНК плазмиды Ti в клетки растений, в результате чего образуется коронный галл (опухоль). Helicobacter pylori использует его для доставки CagA в эпителиальные клетки желудка, чтобы вызвать рак желудка. Bordetella pertussis, бактерия, вызывающая коклюш, выделяет свои токсин коклюша частично через T4SS. Legionella pneumophila, вызывающая легионеллез (болезнь легионеров), имеет T4SS, называемый icm / dot (i ntra c ellular m ultiplication / d воздействуют на o rganelle t гены раффикинга), которые транспортируют многие бактериальные белки в его эукариотического хозяина. Совсем недавно было показано, что фитопатоген Xanthomonas citri использует свой T4SS для секреции эффекторов, летальных для других видов бактерий, что делает эту систему основным детерминантом приспособляемости межвидовой бактериальной конкуренции. Прототипом системы секреции типа IVA является комплекс VirB Agrobacterium tumefaciens.
Системы секреции типа V (T5SS) отличаются от других систем секреции тем, что они секретируют сами себя и вовлекают только внешние клеточная мембрана. Для прохождения секретируемого белка через внутреннюю клеточную мембрану T5SS зависит от системы Sec. У них есть β-бочкообразный домен, который вставляется во внешнюю клеточную мембрану и образует канал, по которому вместе с ним может транспортироваться секретируемый белок. Для этой деятельности их также называют системами автовозов. Когда секретируемые белки выставляются наружу, аутотранспортеры отсекаются (расщепляются), высвобождая белок из домена β-бочонка. Примером автотранспорта является тримерный автотранспортер адгезинов.
Системы секреции типа VI (T6SS) были обнаружены командой Джона Мекаланоса в Гарвардской медицинской школе в г. 2006 из Vibrio cholerae и Pseudomonas aeruginosa. Они были обнаружены, когда мутации в генах Vibrio Cholerae Hcp и VrgG вызвали снижение вирулентности и патогенности. Помимо своей классической роли фактора патогенности, T6SS также участвует в защите от простых эукариотических хищников и во взаимодействиях между бактериями. Ген T6SS образует кластер генов, состоящий из более чем 15 генов. Гены Hcp и VgrG - самые универсальные гены. Структурное сходство T6SS с хвостовым шипом фага T4 позволяет предположить, что процесс инфицирования аналогичен процессу фага.
Система секреции типа VII (T7SS) присутствует в Грамположительные бактерии и микобактерии, такие как M. tuberculosis, M. bovis, Streptomyces coelicolor и S. aureus. В Bacillus subtilis и S. aureus она также называется системой T7b. Он состоит из двух основных компонентов: мембраносвязанной гексамерной АТФазы, которая является членом семейства белков FtsK / SpoIIIE, и любого из белков, связанных с EsxA / EsxB, таких как EsaA, EsaD, EsxB, EsxD, а также Ess. система (EssA, EssB и EsxC найдены у S. aureus). EsxA и EsxB принадлежат к суперсемейству белков WXG100, которые образуют димерные спиральные шпильки. У S. aureus T7SS секретирует большой токсин под названием EsaD, который является членом ферментов нуклеаз. EsaD обезвреживается (детоксифицируется) во время биосинтеза с помощью своего аналога антитоксина EsaG. Затем комплекс EsaD-EsaG связывается с EsaE. Часть EsaE связывается с EssC, который является ферментом АТФазой комплекса T7SS. Во время секреции EsaG остается в цитоплазме, и только EsaD и EsaE секретируются вместе. Но в некоторых штаммах S. aureus EsaD не продуцируется, а вместо этого образуются две копии EsaG-подобных белков. Это могло бы объяснить возникновение T7SS у непатогенных видов, таких как B. subtilis и S. coelicolor.
Системы секреции типа IX (T9SS) регулярно обнаруживаются в Линия бактерий Fibrobacteres-Chlorobi-Bacteroidetes, в которой виды-члены включают внешнюю мембрану. Система по-разному участвует в одном типе скользящей подвижности, в правильном нацеливании определенных факторов вирулентности на поверхность клетки и в разложении комплекса биополимеров. T9SS также известен как секреция Por после орального возбудителя Porphyromonas gingivalis. Описано не менее шестнадцати структурных компонентов системы, включая PorU, транспептидазу, сортирующую белки, которая удаляет C-концевой сигнал сортировки из грузовых белков и вместо этого опосредует их прикрепление к липополисахариду.