
Войти
Слияние везикул - это слияние везикулы с другими везикулами или частью клеточная мембрана. В последнем случае это конечная стадия секреции из секреторных пузырьков, когда их содержимое выводится из клетки посредством экзоцитоза. Везикулы также могут сливаться с другими компартментами клеток-мишеней, такими как лизосома. Экзоцитоз возникает, когда секреторные везикулы временно стыкуются и сливаются в основании чашеобразных структур на плазматической мембране клетки, называемой поросомой, универсальным секреторным механизмом в клетках. Слияние везикул может зависеть от белков SNARE в присутствии повышенной внутриклеточной концентрации кальция (Ca).
Стимулы, запускающие слияние везикул, действуют за счет увеличения внутриклеточного Ca.
Модельные системы, состоящие из одного фосфолипида или его смеси, были изучены физиками-химиками. Кардиолипин содержится в основном в митохондриальных мембранах, а ионы кальция играют важную роль в дыхательных процессах, опосредованных Митохондрия. Постулируемые силы объясняют этот процесс с точки зрения зародышеобразования для агломерации более мелких супрамолекулярных образований или фазовых изменений в структуре биомембран.
При слиянии синаптических пузырьков везикула должна находиться в пределах нескольких нанометров от мембраны-мишени, чтобы начался процесс слияния. Эта близость позволяет клеточной мембране и везикула для обмена липидов, который опосредуется certai n белки, которые удаляют воду, которая попадает между образующимися стыками. Как только везикула находится в нужном положении, она должна ждать, пока Са не войдет в клетку посредством распространения потенциала действия на пресинаптическую мембрану. Са связывается со специфическими белками, одним из которых является синаптотагмин, в нейронах, которые запускают полное слияние везикулы с целевой мембраной.
Также считается, что белки SNARE помогают определять, какая мембрана
 Молекулярный механизм, управляющий экзоцитозом при высвобождении нейромедиатора. Основной комплекс SNARE образован четырьмя α-спиралями, создаваемыми синаптобревином, синтаксином и SNAP-25, синаптотагмин служит сенсором кальция и тесно регулирует застегивание SNARE.
Молекулярный механизм, управляющий экзоцитозом при высвобождении нейромедиатора. Основной комплекс SNARE образован четырьмя α-спиралями, создаваемыми синаптобревином, синтаксином и SNAP-25, синаптотагмин служит сенсором кальция и тесно регулирует застегивание SNARE. Сборка SNARE в «транс» -комплексы, вероятно, связывает мосты между собой. противодействие липидным бислоям мембран, принадлежащих клетке и секреторной грануле, сближая их и вызывая их слияние. Приток кальция в клетку запускает завершение реакции сборки, которая опосредуется взаимодействием между предполагаемым датчиком кальция, синаптотагмином, с липидами мембраны и / или частично собранным комплексом SNARE.
Одна гипотеза предполагает, что молекула Комплексина входит в комплекс SNARE и ее взаимодействие с молекулой синаптотагмина. Известная как гипотеза «зажима», присутствие комплексина обычно ингибирует слияние везикулы с клеточной мембраной. Однако связывание ионов кальция с синаптотагмином вызывает высвобождение или инактивацию комплексина, в результате чего везикула может свободно сливаться.
Согласно гипотезе «застежки-молнии», сборка комплекса начинается с N-конца части мотивов SNARE и продвигается к С-концам, которые закрепляют взаимодействующие белки в мембранах. Формирование комплекса «транс» -SNARE происходит через промежуточный комплекс, состоящий из SNAP-25 и синтаксина-1, который позже вмещает синаптобревин-2 (цитируемые изотипы синтаксина и синаптобревина участвуют в высвобождении нейромедиатора нейронов).
Основываясь на стабильности образующегося комплекса цис-SNARE, было постулировано, что энергия, выделяемая во время процесса сборки, служит средством для преодоления сил отталкивания между мембранами. Есть несколько моделей, которые предлагают объяснение последующей стадии - формирование стебля и поры слияния, но точный характер этих процессов остается обсуждается. Двумя наиболее известными моделями образования поры слияния являются теории пор слияния с липидной и белковой оболочкой.
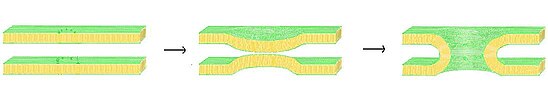 В порах с липидной оболочкой Согласно теории, обе мембраны изгибаются друг к другу, образуя поры раннего слияния. Когда две мембраны находятся на «критическом» расстоянии, липидные головные группы из одной мембраны вставляются в другую, создавая основу для поры слияния.
В порах с липидной оболочкой Согласно теории, обе мембраны изгибаются друг к другу, образуя поры раннего слияния. Когда две мембраны находятся на «критическом» расстоянии, липидные головные группы из одной мембраны вставляются в другую, создавая основу для поры слияния. Одной из возможных моделей образования пор слияния является поры липидных линий. теория. В этой модели, как только мембраны оказываются достаточно близко друг к другу посредством механизма «застежки-молнии» комплекса SNARE, слияние мембран происходит спонтанно. Было показано, что когда две мембраны находятся на критическом расстоянии, гидрофильные липидные головные группы одной мембраны могут сливаться с противоположной мембраной. В модели поры слияния, покрытой липидами, комплекс SNARE действует как каркас, натягивая мембрану, заставляя обе мембраны сморщиваться, чтобы они могли достичь критического расстояния слияния. Когда две мембраны начинают сливаться, образуется покрытый липидами стержень, расширяющийся радиально наружу по мере слияния.
Несмотря на то, что поры, покрытые липидами, возможны и могут обеспечивать все те же свойства, которые наблюдаются при раннем порообразовании, не существует достаточных данных, чтобы доказать, что это единственный метод образования. В настоящее время не существует предложенного механизма межклеточной регуляции флуктуации пор, выстланных липидами, и им было бы значительно труднее производить эффекты, такие как «поцелуй и беги», по сравнению с их аналогами, выстланными белками. Эффективность пор, выстланных липидами, также будет в значительной степени зависеть от состава обеих мембран, и ее успех или неудача могут сильно варьироваться в зависимости от изменений эластичности и жесткости.
Другой возможной моделью образования пор слияния является теория пор с белками. В этой модели после активации синаптотагмина кальцием несколько комплексов SNARE объединяются, чтобы сформировать кольцевую структуру, при этом синаптобревин образует поры в мембране везикул, а синтаксин формирование поры в клеточной мембране. По мере того, как начальная пора расширяется, она включает липиды из обоих бислоев, что в конечном итоге приводит к полному слиянию двух мембран. Комплекс SNARE играет гораздо более активную роль в теории пор, выстланных белками; поскольку поры изначально полностью состоят из белков SNARE, поры легко могут подвергаться межклеточной регуляции, что делает легко достижимыми механизмы флуктуации и «поцелуй и бегство».
Пора, выстланная белками, идеально соответствует всем наблюдаемым требования ранней поры слияния, и хотя некоторые данные подтверждают эту теорию, не существует достаточных данных, чтобы объявить ее основным методом слияния. Для поры, выстланной белком, требуется не менее пяти копий комплекса SNARE, тогда как слияние наблюдалось всего с двумя.
В обеих теориях функция комплекса SNARE остается в основном неизменной, и весь комплекс SNARE остается неизменным. необходимо для начала слияния. Однако было доказано, что in vitro синтаксин сам по себе достаточен для запуска спонтанного кальций-независимого слияния синаптических везикул, содержащих v-SNARE. Это свидетельствует о том, что при Са-зависимом экзоцитозе нейронов синаптотагмин является двойным регулятором, в отсутствие ионов Са он ингибирует динамику SNARE, а в присутствии ионов Са действует как агонист в мембране. процесс слияния.
В синаптических везикулах некоторые нейрохимики предположили, что везикулы иногда могут не полностью сливаться с пресинаптическими мембранами при высвобождении нейромедиатора в синаптическая щель. Разногласия заключаются в том, всегда ли эндоцитоз возникает при реформировании пузырьков после высвобождения нейромедиатора. Другой предложенный механизм высвобождения содержимого везикул во внеклеточную жидкость называется слияние типа «поцелуй и беги».
. Есть некоторые указания на то, что везикулы могут образовывать лишь небольшие поры в пресинаптической мембране, позволяющие высвобождать содержимое путем стандартной диффузии в течение ненадолго, прежде чем отступить обратно в пресинаптическую клетку. Этот механизм может быть способом обхода клатрин-опосредованного эндоцитоза. Также предполагается, что пузырьку не нужно возвращаться в эндосому для пополнения, хотя до конца не известно, с помощью какого механизма он будет заполняться. Это не исключает полного слияния везикул, а только утверждает, что оба механизма могут действовать в синаптических щелях.
Было показано, что «поцелуй и бег» происходит в эндокринных клетках, хотя это не было непосредственно засвидетельствовано в синаптических промежутках.