
Войти
 | |
 | |
| Имена | |
|---|---|
| Другие названия Дифосфопиридин нуклеотид (DPN), Коэнзим I | |
| Идентификаторы | |
| Номер CAS | |
| 3D-модель (JSmol ) | |
| ChEBI | |
| ChEMBL |
|
| ChemSpider | |
| DrugBank | |
| IUPHAR/BPS | |
| KEGG | |
| PubChem CID | |
| номер RTECS |
|
| UNII |
|
InChI
| |
УЛЫБКИ
| |
| Свойства | |
| Химические формула | C21H27N7O14P2 |
| Молярная масса | 663,43 г / моль |
| Внешний вид | Белый порошок |
| Точка плавления | 160 ° C (320 ° F; 433 K) |
| Опасности | |
| Основные опасности | Неопасные |
| NFPA 704 (огненный алмаз) |  1 1 0 1 1 0 |
| Если не указано иное, данные для материалов приводятся в их стандартное состояние (при 25 ° C [77 ° F], 100 кПа). | |
| Ссылки на ink | |
Никотинамидадениндинуклеотид (NAD ) является кофактором, центральным для метаболизма. Обнаруженный во всех живых клетках, НАД называется динуклеотидом, потому что он состоит из двух нуклеотидов, соединенных через их фосфатные группы. Один нуклеотид содержит аденин азотистое основание, а другой никотинамид. НАД существует в двух формах: окисленная и восстановленная форма, сокращенно НАД и НАДН соответственно.
В метаболизме никотинамидадениндинуклеотид участвует в окислительно-восстановительных реакциях, перенося электроны от одной реакции к другой. Таким образом, кофактор присутствует в клетках в двух формах: НАД является окислителем - он принимает электроны от других молекул и становится восстановленным. В результате этой реакции образуется НАДН, который затем может быть использован в качестве восстановителя для передачи электронов. Эти реакции переноса электрона являются основной функцией НАД. Однако он также используется в других клеточных процессах, в первую очередь в качестве субстрата для ферментов при добавлении или удалении химических групп в или из, соответственно, белки, в посттрансляционных модификациях. Из-за важности этих функций ферменты, участвующие в метаболизме НАД, являются мишенями для открытия лекарств.
В организмах НАД может быть синтезирован из простых строительных блоков (de novo ) из любого триптофан или аспарагиновая кислота, каждый из которых представляет собой аминокислоту ; альтернативно, более сложные компоненты коферментов получают из питательных соединений, таких как ниацин ; аналогичные соединения образуются в результате реакций, разрушающих структуру НАД, обеспечивая путь спасения, который «рециркулирует» их обратно в их соответствующую активную форму.
Некоторое количество НАД превращается в кофермент никотинамидадениндинуклеотидфосфат (НАДФ); его химический состав во многом аналогичен химическому составу НАД, хотя преимущественно его роль заключается в качестве кофактора в анаболическом метаболизме.
НАД химические соединения 'добавление с надписями знак отражает формальный заряд на одном из его атомов азота; этот вид представляет собой однозарядный анион - несущий (отрицательный) ионный заряд, равный 1 - в условиях физиологического pH. НАДН, напротив, представляет собой двухзарядный анион из-за его двух мостиковых фосфатных групп.
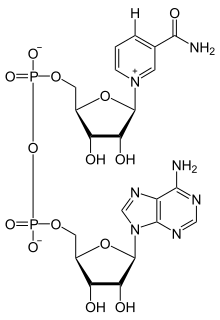
Никотинамидадениндинуклеотид состоит из двух нуклеозидов, соединенных парой мостиковых фосфатных групп. Каждый из нуклеозидов содержит кольцо рибозы, одно с аденином, присоединенным к первому атому углерода (положение 1 ' ), а другое с никотинамидом в этой позиции. Никотинамидная составляющая может быть присоединена к этому аномерному атому углерода в двух ориентациях. Из-за этих двух возможных структур соединение существует в виде двух диастереомеров. Это диастереомер β-никотинамида НАД, который содержится в организмах. Эти нуклеотиды соединены вместе мостиком из двух фосфатных групп через 5'-атомы углерода.
 окислительно-восстановительные реакции никотинамидадениндинуклеотида.
окислительно-восстановительные реакции никотинамидадениндинуклеотида. В метаболизме соединение принимает или отдает электроны в окислительно-восстановительных реакциях. Такие реакции (суммированные в формуле ниже) включают удаление двух атомов водорода из реагента (R) в форме иона гидрида (H) и протона (H). Протон высвобождается в раствор, в то время как восстановитель RH 2 окисляется и НАД восстанавливается до НАДН путем переноса гидрида на никотинамидное кольцо.
Из пары электронов гидрида один электрон переносится на положительно заряженный азот никотинамидного кольца NAD, а второй атом водорода переносится на атом углерода C4 напротив него. азот. потенциал средней точки окислительно-восстановительной пары НАД / НАДН составляет -0,32 вольт, что делает НАДН сильным восстановителем. Реакция легко обратима, когда НАДН восстанавливает другую молекулу и повторно окисляется до НАД. Это означает, что кофермент может непрерывно переключаться между формами НАД и НАДН без потребления.
По внешнему виду все формы этого кофермента представляют собой белые аморфные порошки, которые гигроскопичны и хорошо растворим в воде. Твердые вещества стабильны при хранении в сухом и темном месте. Растворы НАД бесцветны и стабильны примерно в течение недели при 4 ° C и нейтральном pH, но быстро разлагаются в кислотах или щелочах. При разложении они образуют продукты, которые являются ингибиторами ферментов;.
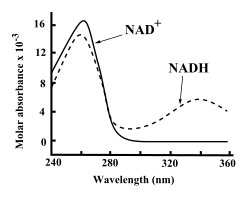 UV спектры поглощения НАД и НАДН.
UV спектры поглощения НАД и НАДН. И НАД, и НАДН сильно поглощают ультрафиолетовый свет из-за аденина. Например, пиковое поглощение НАД находится на длине волны 259 нанометров (нм) с коэффициентом экстинкции 16900 M cm. НАДН также поглощает на более высоких длинах волн, со вторым пиком УФ-поглощения при 339 нм с коэффициентом экстинкции 6220 мкм. Эта разница в ультрафиолетовых спектрах поглощения между окисленной и восстановленной формами коферментов на более высоких длинах волн упрощает измерение превращения одного в другой в ферментных анализах - путем измерения количества УФ-поглощения при 340 нм с использованием спектрофотометра .
НАД и НАДН также различаются по своей флуоресценции. НАДН в растворе имеет пик излучения при 340 нм и время жизни флуоресценции 0,4 наносекунды, в то время как окисленная форма кофермента не флуоресцирует. Свойства сигнала флуоресценции изменяются, когда НАДН связывается с белками, поэтому эти изменения можно использовать для измерения констант диссоциации, которые полезны при изучении кинетики ферментов. Эти изменения флуоресценции также используются для измерения изменений окислительно-восстановительного состояния живых клеток с помощью флуоресцентной микроскопии.
В печени крысы общее количество НАД и НАДН составляет приблизительно 1 мкмоль на грамм сырого веса, что примерно в 10 раз превышает концентрацию НАДФ и НАДФН в тех же клетках. Фактическую концентрацию НАД в клетке цитозоле сложнее измерить: недавние оценки в клетках животных составляют около 0,3 мМ и приблизительно от 1,0 до 2,0 мМ в дрожжах. Однако более 80% флуоресценции НАДН в митохондриях происходит из связанной формы, поэтому концентрация в растворе намного ниже.
Данные для других компартментов клетки ограничены, хотя в митохондрии концентрация НАД аналогична концентрации в цитозоле. Этот НАД переносится в митохондрии с помощью специфического мембранного транспортного белка , поскольку кофермент не может диффундировать через мембраны.
Баланс между окисленной и восстановленной формами никотинамида адениндинуклеотид называется соотношением НАД / НАДН. Это соотношение является важным компонентом того, что называется окислительно-восстановительным состоянием клетки, измерением, которое отражает как метаболическую активность, так и здоровье клеток. Эффекты отношения НАД / НАДН являются сложными, контролируя активность нескольких ключевых ферментов, включая глицеральдегид-3-фосфатдегидрогеназу и пируватдегидрогеназу. В здоровых тканях млекопитающих оценка соотношения между свободным НАД и НАДН в цитоплазме обычно составляет около 700: 1; соотношение, таким образом, благоприятно для окислительных реакций. Отношение общего НАД / НАДН намного ниже, у млекопитающих оценивается от 3 до 10. Напротив, соотношение НАДФ / НАДФН обычно составляет около 0,005, поэтому НАДФН является доминирующей формой этого кофермента. Эти разные соотношения являются ключевыми для различных метаболических ролей НАДН и НАДФН.
НАД синтезируется двумя метаболическими путями. Он продуцируется либо путем de novo из аминокислот, либо путем рециркуляции предварительно сформированных компонентов, таких как никотинамид, обратно в НАД. Хотя большинство тканей у млекопитающих синтезируют НАД путем спасения, гораздо больше de novo синтеза происходит в печени из триптофана, а в почках и макрофагах из никотиновой кислоты.
 Некоторые метаболические пути, которые синтезируют и потребляют НАД у позвоночных. Аббревиатуры определены в тексте.
Некоторые метаболические пути, которые синтезируют и потребляют НАД у позвоночных. Аббревиатуры определены в тексте. Большинство организмов синтезирует НАД из простых компонентов. Конкретный набор реакций у разных организмов различается, но общей чертой является образование хинолиновой кислоты (QA) из аминокислоты - либо триптофана (Trp) у животных и некоторых бактерий, или аспарагиновая кислота (Asp) у некоторых бактерий и растений. Хинолиновая кислота превращается в мононуклеотид никотиновой кислоты (NaMN) путем переноса фрагмента фосфорибозы. Затем аденилатный фрагмент переносится с образованием адениндинуклеотида никотиновой кислоты (NaAD). Наконец, фрагмент никотиновой кислоты в NaAD амидирован до фрагмента никотинамида (Nam) с образованием никотинамидадениндинуклеотида.
На следующем этапе часть NAD превращается в NADP посредством Киназа НАД, которая фосфорилирует НАД. У большинства организмов этот фермент использует АТФ в качестве источника фосфатной группы, хотя некоторые бактерии, такие как Mycobacterium tuberculosis и гипертермофильный архей Pyrococcus horikoshii, используют неорганические полифосфат в качестве альтернативного донора фосфорила.
 Спасательные пути используют три прекурсора НАД.
Спасательные пути используют три прекурсора НАД. Несмотря на наличие пути de novo, спасательные реакции важны в люди; недостаток ниацина в рационе вызывает авитаминоз болезнь пеллагра. Эта высокая потребность в НАД является результатом постоянного потребления кофермента в таких реакциях, как посттрансляционные модификации, поскольку циклическое переключение НАД между окисленной и восстановленной формами в окислительно-восстановительных реакциях не меняет общих уровней кофермента. Основным источником НАД у млекопитающих является спасательный путь, в котором рециклируется никотинамид, продуцируемый ферментами, использующими НАД. Первым этапом и ферментом, ограничивающим скорость в пути спасения, является никотинамидфосфорибозилтрансфераза (NAMPT), которая производит никотинамидмононуклеотид (NMN).
Помимо сборки NAD de novo из простых предшественников аминокислот клетки также утилизируют предварительно образованные соединения, содержащие пиридиновое основание. Три предшественника витаминов, используемые в этих спасательных путях метаболизма, - это никотиновая кислота (NA), никотинамид (Nam) и никотинамидрибозид (NR). Эти соединения могут быть взяты из рациона и называются витамином B 3 или ниацином. Однако эти соединения также производятся внутри клеток и при переваривании клеточного НАД. Некоторые из ферментов, участвующих в этих путях спасения, по-видимому, сконцентрированы в клеточном ядре, что может компенсировать высокий уровень реакций, потребляющих НАД в этой органелле. Имеются сообщения о том, что клетки млекопитающих могут поглощать внеклеточный НАД из своего окружения, и никотинамид и никотинамид рибозид могут абсорбироваться из кишечника.
Пути спасения, используемые в микроорганизмах, отличаются от таковых из млекопитающих. Некоторые патогены, такие как дрожжи Candida glabrata и бактерии Haemophilus influenzae, являются NAD ауксотрофами - они не могут синтезировать NAD - но обладают путями спасения и, таким образом, зависят от внешние источники НАД или его прекурсоров. Еще более удивительным является внутриклеточный патоген Chlamydia trachomatis, в котором отсутствуют распознаваемые кандидаты для каких-либо генов, участвующих в биосинтезе или спасении как НАД, так и НАДФ, и который должен получать эти коферменты из своих хозяин.
 Россманн складывает в части лактатдегидрогеназы из Cryptosporidium parvum, показывая НАД красным, бета-листы желтым и альфа-спирали в фиолетовый.
Россманн складывает в части лактатдегидрогеназы из Cryptosporidium parvum, показывая НАД красным, бета-листы желтым и альфа-спирали в фиолетовый. Никотинамидадениндинуклеотид играет несколько важных ролей в метаболизме. Он действует как кофермент в окислительно-восстановительных реакциях, как донор фрагментов АДФ-рибозы в реакциях АДФ-рибозилирования, как предшественник секунды молекула-мессенджер циклическая АДФ-рибоза, а также действует как субстрат для бактериальных ДНК-лигаз и группы ферментов, называемых сиртуинами, которые используют НАД для удалить ацетильные группы из белков. В дополнение к этим метаболическим функциям НАД представляет собой адениновый нуклеотид, который может высвобождаться из клеток спонтанно и регулируемыми механизмами, и, следовательно, может иметь важные внеклеточные роли.
Основная роль НАД в метаболизме - это перенос электронов от одной молекулы к другой. Реакции этого типа катализируются большой группой ферментов, называемых оксидоредуктазами. Правильные названия этих ферментов содержат названия обоих их субстратов: например, НАДН-убихинон оксидоредуктаза катализирует окисление НАДН коферментом Q. Однако эти ферменты также называют дегидрогеназами или редуктазами, причем НАДН-убихинон оксидоредуктаза обычно называется НАДН-дегидрогеназой или иногда коферментом Q-редуктазой.
Существует множество различных суперсемейств ферментов, которые связывают НАД / НАДН. Одно из наиболее распространенных суперсемейств включает структурный мотив , известный как складка Россмана. Мотив назван в честь Майкла Россманна, который был первым ученым, заметившим, насколько распространена эта структура в нуклеотид-связывающих белках.
Пример бактериального фермента, связывающего НАД, участвующего в метаболизм аминокислот, не имеющий складки Россмана, обнаружен в Pseudomonas syringae pv. помидор (PDB : 2CWH ; InterPro : IPR003767 ).
 На этой диаграмме акцептор гидрида углерода C4 показан вверху. никотинамидное кольцо лежит в плоскости страницы с карбоксиамидом справа, как показано, донор гидрида находится либо «выше», либо «ниже» плоскости страницы. Если «выше» перенос гидрида относится к классу A, если «нижний» перенос гидрида относится к классу B.
На этой диаграмме акцептор гидрида углерода C4 показан вверху. никотинамидное кольцо лежит в плоскости страницы с карбоксиамидом справа, как показано, донор гидрида находится либо «выше», либо «ниже» плоскости страницы. Если «выше» перенос гидрида относится к классу A, если «нижний» перенос гидрида относится к классу B. При связывании в активном центре оксидоредуктазы никотинамидное кольцо кофермента располагается так, что оно может принимать гидрид от другого субстрата. В зависимости от фермента гидрид донор расположен либо «над», либо «под» плоскостью плоского углерода C4, как показано на рисунке. Оксидоредуктазы класса A переносят атом сверху; ферменты класса B переносят его снизу. Поскольку углерод C4, который принимает водород является прохиральным, его можно использовать в кинетике фермента для получения информации о механизме действия фермента. Это достигается путем смешивания фермент с субстратом, у которого атомы водорода замещены, поэтому фермент будет восстанавливать НАД путем переноса дейтерия, а не водорода. В этом случае фермент может продуцировать один из двух стереоизомеров НАДН.
Несмотря на сходство в том, как белки связывают два кофермента, ферменты почти всегда демонстрируют высокий уровень специфичности для любого НАД. или НАДФ. Эта специфичность отражает различные метаболические роли соответствующих коферментов и является результатом различных наборов аминокислотных остатков в двух типах кофермент-связывающего кармана. Например, в активном центре НАДФ-зависимых ферментов между основной боковой цепью аминокислоты и кислой фосфатной группой НАДФ образуется ионная связь . Напротив, в НАД-зависимых ферментах заряд в этом кармане меняется на противоположный, предотвращая связывание НАДФ. Однако есть несколько исключений из этого общего правила, и такие ферменты, как альдозоредуктаза, глюкозо-6-фосфатдегидрогеназа и метилентетрагидрофолатредуктаза, могут использовать оба коферменты у некоторых видов.
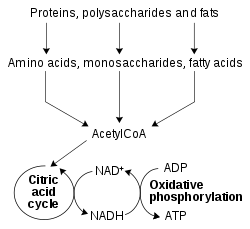 Упрощенная схема окислительно-восстановительного метаболизма, показывающая, как НАД и НАДН связывают цикл лимонной кислоты и окислительное фосфорилирование.
Упрощенная схема окислительно-восстановительного метаболизма, показывающая, как НАД и НАДН связывают цикл лимонной кислоты и окислительное фосфорилирование.Окислительно-восстановительные реакции, катализируемые оксидоредуктазами, жизненно важны для всех звеньев метаболизма, но одна особенно важная функция этих реакций - позволить питательным веществам разблокировать энергию, хранящуюся в относительно слабой двойной связи кислорода. Здесь восстановленные соединения, такие как глюкоза и жирные кислоты, окисляются, тем самым высвобождая химическую энергию O 2. В этом процессе НАД восстанавливается до НАДН как часть бета-окисления, гликолиза и цикла лимонной кислоты. У эукариот электроны, переносимые НАДН, который продуцируется в цитоплазме, переносятся в митохондрию (для уменьшения митохондриального НАД) митохондриальными шаттлами, например, челнок малат-аспартат. Затем митохондриальный НАДН окисляется, в свою очередь, с помощью цепи переноса электронов, которая перекачивает протоны через мембрану и генерирует АТФ посредством окислительного фосфорилирования. Эти челночные системы также выполняют ту же транспортную функцию в хлоропластах.
Поскольку в этих связанных наборах реакций используются как окисленная, так и восстановленная формы никотинамидадениндинуклеотида, клетка поддерживает значительные концентрации как NAD, так и NADH, с высокое соотношение НАД / НАДН, позволяющее этому коферменту действовать как окислитель и как восстановитель. Напротив, основная функция НАДФН - это восстанавливающий агент в анаболизме, при этом этот кофермент участвует в таких путях, как синтез жирных кислот и фотосинтез. Поскольку НАДФН необходим для запуска окислительно-восстановительных реакций в качестве сильного восстановителя, соотношение НАДФ / НАДФН поддерживается на очень низком уровне.
Хотя он важен для катаболизма, НАДН также используется в анаболических реакциях, таких как глюконеогенез. Эта потребность в НАДН в анаболизме создает проблему для прокариот, растущих на питательных веществах, выделяющих лишь небольшое количество энергии. Например, нитрифицирующие бактерии, такие как Nitrobacter, окисляют нитрит до нитрата, который выделяет достаточно энергии, чтобы перекачивать протоны и генерировать АТФ, но недостаточно для прямого производства NADH. Поскольку НАДН по-прежнему необходим для анаболических реакций, эти бактерии используют нитрит-оксидоредуктазу для выработки протонодвижущей силы, достаточной для запуска части цепи переноса электронов в обратном направлении, генерируя НАДН.
Кофермент НАД также расходуется в реакциях переноса АДФ-рибозы. Например, ферменты, называемые АДФ-рибозилтрансферазы, добавляют фрагмент АДФ-рибозы этой молекулы к белкам в посттрансляционной модификации, называемой АДФ-рибозилированием. АДФ-рибозилирование включает либо добавление одной части АДФ-рибозы при моно-АДФ-рибозилировании, либо перенос АДФ-рибозы к белкам в длинных разветвленных цепях, что называется поли (АДФ-рибозилирование). Моно-АДФ-рибозилирование было впервые идентифицировано как механизм группы бактериальных токсинов, особенно холерного токсина, но оно также участвует в передаче сигналов нормальных клеток. Поли (АДФ-рибозилирование) осуществляется полимеразами поли (АДФ-рибоза). Структура поли (АДФ-рибоза) участвует в регуляции нескольких клеточных событий и наиболее важна в ядре клетки, в таких процессах, как репарация ДНК и теломер обслуживание. В дополнение к этим функциям внутри клетки недавно была обнаружена группа внеклеточных АДФ-рибозилтрансфераз, но их функции остаются неясными. НАД также может быть добавлен к клеточной РНК в виде 5'-концевой модификации.
 Структура циклической АДФ-рибозы.
Структура циклической АДФ-рибозы.Другой функцией этого кофермента в передаче сигналов в клетке является его предшественник. циклической АДФ-рибозы, которая продуцируется из НАД с помощью АДФ-рибозилциклазы, как часть системы второго мессенджера. Эта молекула действует в передаче сигналов кальция, высвобождая кальций из внутриклеточных запасов. Он делает это путем связывания и открытия класса кальциевых каналов, называемых рианодиновых рецепторов, которые расположены в мембранах органелл, таких как эндоплазматический ретикулум.
NAD. также потребляется сиртуинами, которые представляют собой НАД-зависимые деацетилазы, такие как Sir2. Эти ферменты действуют путем переноса ацетильной группы со своего субстратного белка на АДФ-рибозный фрагмент НАД; это расщепляет кофермент и высвобождает никотинамид и О-ацетил-АДФ-рибозу. Сиртуины, по-видимому, в основном участвуют в регуляции транскрипции посредством деацетилирования гистонов и изменения структуры нуклеосомы. Однако сиртуины также могут деацетилировать негистоновые белки. Эти активности сиртуинов особенно интересны из-за их важности в регуляции старения.
Другие НАД-зависимые ферменты включают бактериальные ДНК-лигазы, которые соединяют два конца ДНК, используя НАД в качестве субстрата для отдать часть аденозинмонофосфата (AMP) 5'-фосфату одного конца ДНК. Затем это промежуточное соединение атакует 3'-гидроксильная группа другого конца ДНК, образуя новую фосфодиэфирную связь. Это контрастирует с эукариотическими ДНК-лигазами, которые используют АТФ для образования промежуточного соединения ДНК-АМФ.
Li et al. обнаружили, что НАД напрямую регулирует межбелковые взаимодействия. Они также показывают, что одной из причин возрастного снижения репарации ДНК может быть повышенное связывание белка DBC1 (удалено при раке молочной железы 1) с PARP1 (поли [ADP– рибоза] полимераза 1), поскольку уровни НАД снижаются во время старения. Таким образом, модуляция НАД может защищать от рака, радиации и старения.
В последние годы НАД также был признан внеклеточным сигнальная молекула, участвующая в межклеточной коммуникации. НАД высвобождается из нейронов в кровеносных сосудах, мочевом пузыре, толстой кишке, из нейросекреторных клеток и из синаптосом мозга и предлагается в качестве нового нейромедиатора, который передает информацию от нервов эффекторным клеткам в гладкомышечных органах. У растений внеклеточный никотинамидадениндинуклеотид индуцирует устойчивость к инфекции патогена, и был идентифицирован первый внеклеточный рецептор NAD. Необходимы дальнейшие исследования, чтобы определить основные механизмы его внеклеточного действия и их важность для здоровья человека и жизненных процессов в других организмах.
Ферменты, которые производят и используют НАД и НАДН, важны как для фармакологии, так и для исследования будущих методов лечения заболеваний. Дизайн лекарств и разработка лекарств использует НАД тремя способами: как прямую мишень для лекарств, путем создания ингибиторов ферментов или активаторов на основе его структуры, которые изменяют активность НАД-зависимых ферментов, и путем попытки ингибировать биосинтез НАД..
Поскольку раковые клетки используют повышенный гликолиз, и поскольку НАД усиливает гликолиз, никотинамидфосфорибозилтрансфераза (путь восстановления НАД) часто усиливается в раковых клетках.
Это было изучено для его потенциального использования в терапии нейродегенеративных заболеваний, таких как болезнь Альцгеймера и болезнь Паркинсона. Плацебо-контролируемое клиническое испытание НАДН (которое исключало предшественники НАДН) у людей с болезнью Паркинсона не продемонстрировало какого-либо эффекта.
НАД также является прямой мишенью лекарственного средства изониазид, который используется для лечения туберкулеза, инфекции, вызываемой Mycobacterium tuberculosis. Изониазид представляет собой пролекарство , и как только оно попадает в бактерии, оно активируется ферментом пероксидазой, который окисляет соединение до формы свободных радикалов. Затем этот радикал вступает в реакцию с NADH с образованием аддуктов, которые являются очень мощными ингибиторами ферментов еноилацил-протеин-редуктаза и дигидрофолатредуктазы.
, поскольку большое количество оксидоредуктаз используют NAD и NADH. в качестве субстратов и связывают их с помощью высококонсервативного структурного мотива, идея о том, что ингибиторы на основе NAD могут быть специфичными для одного фермента, является неожиданной. Однако это возможно: например, ингибиторы на основе соединений микофеноловой кислоты и тиазофурина ингибируют IMP-дегидрогеназу в сайте связывания NAD. Из-за важности этого фермента в метаболизме пуринов эти соединения могут быть полезны в качестве противораковых, противовирусных или иммунодепрессантов. Другие препараты не являются ингибиторами ферментов, а вместо этого активируют ферменты, участвующие в метаболизме НАД. Сиртуины являются особенно интересной мишенью для таких лекарств, поскольку активация этих НАД-зависимых деацетилаз увеличивает продолжительность жизни в некоторых моделях животных. Соединения, такие как ресвератрол, повышают активность этих ферментов, что может иметь важное значение для их способности замедлять старение как у позвоночных, так и у беспозвоночных модельных организмов. В одном эксперименте у мышей, получавших НАД в течение одной недели, улучшилась ядерно-митохрондриальная связь.
Из-за различий в метаболических путях биосинтеза НАД между организмами, например между бактериями и людьми, Эта область метаболизма является многообещающей для разработки новых антибиотиков. Например, фермент никотинамидаза, который превращает никотинамид в никотиновую кислоту, является мишенью для разработки лекарств, поскольку этот фермент отсутствует у людей, но присутствует в дрожжах и бактериях.
В бактериологии, НАД, иногда называемый фактором V, используется как добавка к питательной среде для некоторых привередливых бактерий.
 Артур Харден, соавтор обнаружения NAD
Артур Харден, соавтор обнаружения NAD Коэнзим НАД был впервые открыт британскими биохимиками Артуром Харденом и Уильямом Джоном Янгом в 1906 году. Они заметили, что добавление вареных и фильтрованных дрожжей Экстракт сильно ускоренного спиртового брожения в некипяченых дрожжевых экстрактах. Они назвали неопознанный фактор, ответственный за этот эффект, коферментом. Путем длительной и сложной очистки от дрожжевых экстрактов этот термостабильный фактор был идентифицирован Гансом фон Эйлер-Челпином как нуклеотид фосфат сахара. В 1936 году немецкий ученый Отто Генрих Варбург показал функцию нуклеотидного кофермента в переносе гидрида и идентифицировал никотинамидную часть как место окислительно-восстановительных реакций.
Витамин предшественники НАД были впервые идентифицированы в 1938 году, когда Конрад Эльвехджем показал, что печень обладает активностью «против черного языка» в форме никотинамида. Затем, в 1939 году, он представил первые убедительные доказательства того, что ниацин используется для синтеза НАД. В начале 1940-х годов Артур Корнберг первым обнаружил фермент в пути биосинтеза. В 1949 г. американские биохимики Моррис Фридкин и Альберт Л. Ленингер доказали, что НАДН связывает метаболические пути, такие как цикл лимонной кислоты, с синтезом АТФ в окислительной фосфорилирование. В 1958 году Джек Прейсс и Филип Хэндлер открыли промежуточные соединения и ферменты, участвующие в биосинтезе НАД; вторичный синтез из никотиновой кислоты называется путем Прейсс-Хэндлера. В 2004 году Чарльз Бреннер и его сотрудники раскрыли никотинамидрибозид киназный путь к NAD.
Не-окислительно-восстановительные роли NAD (P) были обнаружены позже. Первым, что было выявлено, было использование НАД в качестве донора АДФ-рибозы в реакциях АДФ-рибозилирования, наблюдавшихся в начале 1960-х годов. Исследования 1980-х и 1990-х годов выявили активность НАД и метаболитов НАДФ в передаче сигналов в клетках, такую как действие циклической АДФ-рибозы, которое было обнаружено в 1987 году. Метаболизм НАД оставался областью интенсивных исследований. в 21 век, с повышенным интересом после открытия NAD-зависимых протеин-деацетилаз, названных сиртуинами в 2000 году, Шин-ичиро Имаи и его коллеги.
| На Викискладе есть средства массовой информации, связанные с Никотинамидадениндинуклеотид. |