
Войти
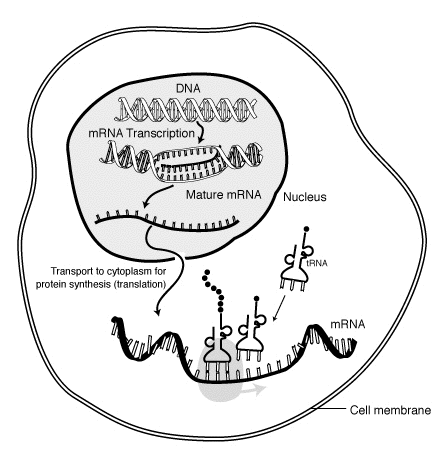 «Жизненный цикл» мРНК в эукариотической клетке. РНК является транскрибируется в ядре ; после обработки, он транспортируется в цитоплазму и переведен с помощью рибосом. Наконец, мРНК деградирует.
«Жизненный цикл» мРНК в эукариотической клетке. РНК является транскрибируется в ядре ; после обработки, он транспортируется в цитоплазму и переведен с помощью рибосом. Наконец, мРНК деградирует. В молекулярной биологии, мессенджер рибонуклеиновая кислота ( мРНК) представляет собой одноцепочечная молекула из РНКА, которая соответствует генетической последовательности о наличии гена, и считываются рибосомой в процессе синтеза в белках.
мРНК создается в процессе транскрипции, когда фермент ( РНК-полимераза ) преобразует ген в мРНК первичного транскрипта (также известную как пре-мРНК ). Эта пре-мРНК обычно все еще содержит интроны, участки, которые не будут кодировать окончательную аминокислотную последовательность. Они удаляются в процессе сплайсинга РНК, оставляя только экзоны, участки, которые будут кодировать белок. Эта последовательность экзона составляет зрелую мРНК. Затем зрелая мРНК считывается рибосомой, и, используя аминокислоты, переносимые транспортной РНК (тРНК), рибосома создает белок. Этот процесс известен как перевод. Все эти процессы составляют часть центральной догмы молекулярной биологии, которая описывает поток генетической информации в биологической системе.
Как и в ДНК, генетическая информация в мРНК содержится в последовательности нуклеотидов, которые организованы в кодоны, каждый из которых состоит из трех рибонуклеотидов. Каждый кодон кодирует определенную аминокислоту, за исключением стоп-кодонов, которые прекращают синтез белка. Для трансляции кодонов в аминокислоты требуются два других типа РНК: транспортная РНК, которая распознает кодон и обеспечивает соответствующую аминокислоту, и рибосомная РНК (рРНК), центральный компонент механизма производства белка рибосом.
Идея мРНК была впервые предложена Сиднеем Бреннером и Фрэнсисом Криком 15 апреля 1960 года в Королевском колледже в Кембридже, когда Франсуа Жакоб рассказывал им о недавнем эксперименте, проведенном самим Артуром Парди и Жаком Моно. При поддержке Крика Бреннер и Джейкоб немедленно приступили к проверке этой новой гипотезы и обратились к Мэтью Мезельсону из Калифорнийского технологического института. Летом 1960 года Бреннер, Джейкоб и Мезельсон провели эксперимент в лаборатории Мезельсона в Калифорнийском технологическом институте, который установил существование мРНК. Той осенью Джейкоб и Моно придумали название «РНК-мессенджер» и разработали первую теоретическую основу для объяснения ее функции. В феврале 1961 года Джеймс Уотсон обнаружил, что его исследовательская группа идет за ними с аналогичным экспериментом примерно в том же направлении; Бреннер и другие согласились на просьбу Уотсона отложить публикацию результатов их исследования. В результате статьи Бреннера и Уотсона были опубликованы одновременно в том же номере журнала Nature в мае 1961 года, а в том же месяце Джейкоб и Моно опубликовали свою теоретическую основу для мРНК в Journal of Molecular Biology.
Кратковременное существование молекулы мРНК начинается с транскрипции и в конечном итоге заканчивается деградацией. В течение своей жизни молекула мРНК также может обрабатываться, редактироваться и транспортироваться перед трансляцией. Молекулы мРНК эукариот часто требуют обширной обработки и транспорта, в то время как молекулы мРНК прокариот - нет. Молекула мРНК эукариот и окружающие ее белки вместе называются РНП-мессенджером.
Транскрипция - это когда РНК копируется из ДНК. Во время транскрипции РНК-полимераза при необходимости копирует ген из ДНК в мРНК. Этот процесс немного отличается у эукариот и прокариот. Одно заметное отличие состоит в том, что прокариотическая РНК-полимераза связывается с ферментами, обрабатывающими ДНК, во время транскрипции, так что процессинг может происходить во время транскрипции. Следовательно, это приводит к тому, что новая цепь мРНК становится двухцепочечной, образуя комплементарную цепь, известную как цепь тРНК, которые при объединении не могут образовывать структуры из спаривания оснований. Более того, матрицей для мРНК является комплементарная цепь тРНК, которая идентична по последовательности антикодоновой последовательности, с которой связывается ДНК. Короткоживущий, необработанный или частично процессированный продукт называется мРНК-предшественником или пре-мРНК ; после полной обработки она называется зрелой мРНК.
Обработка мРНК сильно различается у эукариот, бактерий и архей. Неэукариотическая мРНК, по сути, созревает после транскрипции и не требует обработки, за исключением редких случаев. Однако эукариотическая пре-мРНК требует нескольких этапов обработки, прежде чем она будет доставлена в цитоплазму и ее трансляция рибосомой.
Обширный процессинг эукариотической пре-мРНК, который приводит к зрелой мРНК, представляет собой сплайсинг РНК, механизм, с помощью которого интроны или аутроны (некодирующие области) удаляются и экзоны (кодирующие области) соединяются вместе.
5' крышки (также называемая цоколем РНК, РНК - 7-метилгуанозин колпачок, или РНК, м 7 G колпачок) является то, что была добавлена к „фронт“ или модифицированный гуанин нуклеотид 5' - конце эукариотической матричной РНК в скором времени после начала транскрипции. 5'-кэп состоит из концевого остатка 7-метилгуанозина, который связан через 5'-5'-трифосфатную связь с первым транскрибированным нуклеотидом. Его присутствие имеет решающее значение для распознавания рибосомой и защиты от РНКаз.
Добавление кэпа связано с транскрипцией и происходит совместно с транскрипцией, так что одно влияет на другое. Вскоре после начала транскрипции 5'-конец синтезируемой мРНК связывается синтезирующим кэп комплексом, связанным с РНК-полимеразой. Этот ферментный комплекс катализирует химические реакции, необходимые для кэппирования мРНК. Синтез протекает как многоступенчатая биохимическая реакция.
В некоторых случаях мРНК будет редактироваться, изменяя нуклеотидный состав этой мРНК. Примером у людей является мРНК аполипопротеина B, которая редактируется в некоторых тканях, но не редактируется в других. Редактирование создает ранний стоп-кодон, который после трансляции дает более короткий белок.
Полиаденилирование - это ковалентное связывание полиаденилильного фрагмента с молекулой информационной РНК. В эукариотических организмах большинство молекул матричной РНК (мРНК) полиаденилированы на 3'-конце, но недавние исследования показали, что короткие участки уридина (олигоуридилирование) также обычны. Поли (А) хвост и белок связан с ней помочь в защите от деградации мРНК экзонуклеазами. Полиаденилирование также важно для терминации транскрипции, экспорта мРНК из ядра и трансляции. мРНК также может быть полиаденилирована в прокариотических организмах, где поли (A) хвосты действуют скорее для облегчения, чем препятствия экзонуклеолитической деградации.
Полиаденилирование происходит во время и / или сразу после транскрипции ДНК в РНК. После завершения транскрипции цепь мРНК расщепляется под действием эндонуклеазного комплекса, связанного с РНК-полимеразой. После расщепления мРНК к свободному 3'-концу в сайте расщепления добавляется около 250 остатков аденозина. Эта реакция катализируется полиаденилат-полимеразой. Так же, как и при альтернативном сплайсинге, может быть более одного варианта полиаденилирования мРНК.
Встречаются также мутации сайта полиаденилирования. Первичный транскрипт РНК гена расщепляется по сайту присоединения поли-А, и 100-200 А добавляются к 3'-концу РНК. Если этот сайт изменен, образуется аномально длинная и нестабильная конструкция мРНК.
Еще одно различие между эукариотами и прокариотами - транспорт мРНК. Поскольку транскрипция и трансляция эукариот разделены по компартментам, мРНК эукариот должны экспортироваться из ядра в цитоплазму - процесс, который может регулироваться разными сигнальными путями. Зрелые мРНК распознаются посредством их процессированных модификаций и затем экспортируются через ядерную пору за счет связывания с кэп-связывающими белками CBP20 и CBP80, а также с комплексом транскрипции / экспорта (TREX). Множественные пути экспорта мРНК были идентифицированы у эукариот.
В пространственно сложных клетках некоторые мРНК транспортируются в определенные субклеточные пункты назначения. В зрелых нейронах определенные мРНК транспортируются от сомы к дендритам. Один сайт трансляции мРНК находится в полирибосомах, выборочно локализованных под синапсами. МРНК для Arc / Arg3.1 индуцируется синаптической активностью и селективно локализуется вблизи активных синапсов на основе сигналов, генерируемых рецепторами NMDA. Другие мРНК также перемещаются в дендриты в ответ на внешние стимулы, такие как мРНК β-актина. При экспорте из ядра мРНК актина связывается с ZBP1 и субъединицей 40S. Комплекс связывается моторным белком и транспортируется к месту назначения (расширение нейрита) вдоль цитоскелета. В конечном итоге ZBP1 фосфорилируется с помощью Src, чтобы инициировать трансляцию. В развивающихся нейронах мРНК также транспортируются в растущие аксоны и особенно в конусы роста. Многие мРНК помечены так называемыми «почтовыми индексами», которые нацелены на их транспортировку в определенное место.
Поскольку прокариотическая мРНК не нуждается в процессинге или транспортировке, трансляция рибосомой может начаться сразу после окончания транскрипции. Следовательно, можно сказать, что прокариотическая трансляция связана с транскрипцией и происходит котранскрипционно.
Эукариотическая мРНК, которая была обработана и транспортирована в цитоплазму (т.е. зрелая мРНК), затем может транслироваться рибосомой. Перевод может происходить на рибосомах свободно плавающие в цитоплазме, или направленный на эндоплазматический ретикулум посредством частицы распознавания сигнала. Следовательно, в отличие от прокариот, трансляция эукариот напрямую не связана с транскрипцией. В некоторых случаях возможно даже, что снижение уровней мРНК сопровождается повышенными уровнями белка, как это наблюдалось для уровней мРНК / белка EEF1A1 при раке молочной железы.
 Структура зрелой мРНК эукариот. Полностью процессированная мРНК включает 5'-кэп, 5'-UTR, кодирующую область, 3'-UTR и поли (A) хвост.
Структура зрелой мРНК эукариот. Полностью процессированная мРНК включает 5'-кэп, 5'-UTR, кодирующую область, 3'-UTR и поли (A) хвост. Кодирующие области состоят из кодонов, которые декодируются и транслируются в белки рибосомой; у эукариот обычно на одну, а у прокариот обычно на несколько. Кодирующие области начинаются стартовым кодоном и заканчиваются стоп-кодоном. В общем, стартовый кодон представляет собой триплет AUG, а стоп-кодон - UAG («янтарный»), UAA («охра») или UGA («опал»). Кодирующие области обычно стабилизируются внутренними парами оснований, что препятствует деградации. Помимо того, что они кодируют белок, части кодирующих областей могут служить регуляторными последовательностями в пре-мРНК в качестве энхансеров экзонного сплайсинга или сайленсеров экзонного сплайсинга.
Нетранслируемые области (UTR) представляют собой участки мРНК перед стартовым кодоном и после стоп-кодона, которые не транслируются, называемые пятью первичными нетранслируемыми областями (5 'UTR) и тремя первичными нетранслированными областями (3' UTR), соответственно. Эти области транскрибируются с кодирующей областью и, таким образом, являются экзонными, поскольку они присутствуют в зрелой мРНК. Нетранслируемым областям приписывается несколько ролей в экспрессии генов, включая стабильность мРНК, локализацию мРНК и эффективность трансляции. Способность UTR выполнять эти функции зависит от последовательности UTR и может различаться для разных мРНК. Генетические варианты в 3 'UTR также участвуют в восприимчивости к заболеванию из-за изменения структуры РНК и трансляции белка.
Стабильность мРНК можно контролировать с помощью 5 'UTR и / или 3' UTR из-за различного сродства к ферментам, разрушающим РНК, называемым рибонуклеазами, и к вспомогательным белкам, которые могут стимулировать или ингибировать деградацию РНК. (См. Также C-богатый элемент стабильности. )
Эффективность трансляции, включая иногда полное запрещение трансляции, можно контролировать с помощью UTR. Белки, которые связываются с 3 'или 5' UTR, могут влиять на трансляцию, влияя на способность рибосомы связываться с мРНК. МикроРНК, связанные с 3 'UTR, также могут влиять на эффективность трансляции или стабильность мРНК.
Считается, что цитоплазматическая локализация мРНК является функцией 3 'UTR. Белки, которые необходимы в определенной области клетки, также могут транслироваться туда; в таком случае 3'-UTR может содержать последовательности, которые позволяют транскрипту быть локализованным в этой области для трансляции.
Некоторые элементы, содержащиеся в нетранслируемых областях, при транскрибировании в РНК образуют характерную вторичную структуру. Эти структурные элементы мРНК участвуют в регуляции мРНК. Некоторые из них, такие как элемент SECIS, являются мишенями для связывания белков. Один класс элементов мРНК, рибопереключатели, напрямую связывают небольшие молекулы, изменяя их укладку, чтобы изменить уровни транскрипции или трансляции. В этих случаях мРНК регулируется сама.
3'-поли (A) -хвост представляет собой длинную последовательность адениновых нуклеотидов (часто несколько сотен), добавленных к 3'-концу пре-мРНК. Этот хвост способствует экспорту из ядра и трансляции и защищает мРНК от деградации.
Молекула мРНК называется моноцистронной, если она содержит генетическую информацию для трансляции только одной белковой цепи (полипептида). Так обстоит дело с большинством мРНК эукариот. С другой стороны, полицистронная мРНК несет несколько открытых рамок считывания (ORF), каждая из которых транслируется в полипептид. Эти полипептиды обычно имеют родственную функцию (они часто представляют собой субъединицы, составляющие конечный комплексный белок), и их кодирующая последовательность сгруппирована и регулируется вместе в регуляторной области, содержащей промотор и оператор. Большая часть мРНК, обнаруженной у бактерий и архей, является полицистронной, как и митохондриальный геном человека. Дицистронная или бицистронная мРНК кодирует только два белка.
У эукариот молекулы мРНК образуют кольцевые структуры из-за взаимодействия между eIF4E и поли (A) -связывающим белком, которые оба связываются с eIF4G, образуя мостик мРНК-белок-мРНК. Считается, что циркуляризация способствует циклированию рибосом на мРНК, что приводит к эффективной по времени трансляции, а также может обеспечивать транслирование только интактной мРНК (для частично деградированной мРНК характерно отсутствие кэпа m7G или поли-A-хвоста).
Существуют и другие механизмы циркуляризации, особенно в мРНК вируса. В мРНК полиовируса используется участок клеверного листа ближе к 5'-концу для связывания PCBP2, который связывает поли (A) -связывающий белок, образуя знакомый круг мРНК-белок-мРНК. Вирус желтого карлика ячменя имеет связывание между сегментами мРНК на его 5'-конце и 3'-конце (так называемые петли «целующегося стебля»), что приводит к циркуляризации мРНК без участия каких-либо белков.
Геномы РНК-вирусов (+ цепи которых транслируются как мРНК) также обычно подвергаются циркуляризации. Во время репликации генома циркуляризация действует для увеличения скорости репликации генома, циклически повторяя вирусную РНК-зависимую РНК-полимеразу так же, как предполагается, что рибосома циклизуется.
Различные мРНК в одной и той же клетке имеют разное время жизни (стабильность). В бактериальных клетках отдельные мРНК могут выжить от секунд до более часа. Однако время жизни в среднем составляет от 1 до 3 минут, что делает бактериальную мРНК гораздо менее стабильной, чем мРНК эукариот. В клетках млекопитающих время жизни мРНК колеблется от нескольких минут до дней. Чем выше стабильность мРНК, тем больше белка может быть произведено из этой мРНК. Ограниченное время жизни мРНК позволяет клетке быстро изменять синтез белка в ответ на меняющиеся потребности. Существует множество механизмов, которые приводят к разрушению мРНК, некоторые из которых описаны ниже.
В целом у прокариот время жизни мРНК намного короче, чем у эукариот. Прокариоты разрушают сообщения с помощью комбинации рибонуклеаз, включая эндонуклеазы, 3'-экзонуклеазы и 5'-экзонуклеазы. В некоторых случаях малые молекулы РНК (мРНК) длиной от десятков до сотен нуклеотидов могут стимулировать деградацию специфических мРНК за счет спаривания оснований с комплементарными последовательностями и облегчения расщепления рибонуклеазой РНКазой III. Недавно было показано, что бактерии также имеют своего рода 5-футовый колпачок, состоящий из трифосфата на 5'-конце. Удаление двух фосфатов оставляет 5'-монофосфат, в результате чего сообщение разрушается экзонуклеазой РНКазы J, которая разлагается с 5 'до 3'.
Внутри эукариотических клеток существует баланс между процессами трансляции и распада мРНК. Активно транслируемые сообщения связываются рибосомами, факторами инициации эукариот eIF-4E и eIF-4G и поли (A) -связывающим белком. eIF-4E и eIF-4G блокируют расщепляющий фермент ( DCP2 ), а поли (A) -связывающий белок блокирует экзосомный комплекс, защищая концы сообщения. Баланс между трансляцией и распадом отражается в размере и количестве цитоплазматических структур, известных как P-тельца. Поли (A) хвост мРНК укорачивается специализированными экзонуклеазами, которые нацелены на определенные информационные РНК с помощью комбинации цис-регуляторных последовательностей. на РНК и транс-действующих РНК-связывающих белках. Считается, что удаление поли (A) хвоста нарушает круговую структуру сообщения и дестабилизирует комплекс связывания кэпа. Сообщение затем подвергается разложению либо экзосомой или decapping комплекса. Таким образом, трансляционно неактивные сообщения могут быть быстро уничтожены, в то время как активные сообщения остаются нетронутыми. Механизм остановки трансляции и передачи сообщения комплексам распада в деталях не изучен.
Присутствие AU-богатых элементов в некоторых мРНК млекопитающих имеет тенденцию дестабилизировать эти транскрипты за счет действия клеточных белков, которые связывают эти последовательности и стимулируют удаление поли (A) хвоста. Считается, что потеря поли (A) хвоста способствует деградации мРНК, облегчая атаку как экзосомным комплексом, так и комплексом декапирования. Быстрая деградация мРНК с помощью элементов, богатых AU, является критическим механизмом для предотвращения перепроизводства мощных цитокинов, таких как фактор некроза опухоли (TNF) и фактор, стимулирующий колонии гранулоцитов-макрофагов (GM-CSF). AU-богатые элементы также регулируют биосинтез протоонкогенных факторов транскрипции, таких как c-Jun и c-Fos.
Эукариотические сообщения подлежат наблюдению с помощью бессмысленного распада (NMD), который проверяет наличие преждевременных стоп-кодонов (бессмысленных кодонов) в сообщении. Они могут возникать из-за неполного сплайсинга, рекомбинации V (D) J в адаптивной иммунной системе, мутаций в ДНК, ошибок транскрипции, негерметичного сканирования рибосомой, вызывающего сдвиг рамки, и других причин. Обнаружение преждевременного стоп-кодона запускает деградацию мРНК за счет 5 'декапирования, удаления 3' поли (A) хвоста или эндонуклеолитического расщепления.
В многоклеточных, малые интерферирующие РНК (киРНК), обработанные Dicer включены в комплекс, известный как РНК-индуцированного глушителей комплекс или RISC -. Этот комплекс содержит эндонуклеазу, которая расщепляет идеально комплементарные сообщения, с которыми связывается миРНК. Затем полученные фрагменты мРНК разрушаются экзонуклеазами. siRNA обычно используется в лабораториях для блокирования функции генов в культуре клеток. Считается, что он является частью врожденной иммунной системы как защита от вирусов с двухцепочечной РНК.
МикроРНК (миРНК) представляют собой небольшие РНК, которые обычно частично комплементарны последовательностям информационных РНК многоклеточных животных. Связывание miRNA с сообщением может подавлять трансляцию этого сообщения и ускорять удаление поли (A) хвоста, тем самым ускоряя деградацию мРНК. Механизм действия миРНК является предметом активных исследований.
Есть и другие способы, с помощью которых сообщения могут быть разрушены, в том числе безостановочный распад и молчание с помощью Piwi-взаимодействующей РНК (piRNA), среди прочего.
Введение мессенджера РНК нуклеозидов с модифицированной последовательностью может вызвать клетку, чтобы сделать белка, который, в свою очередь, может непосредственно лечить заболевание или могут функционировать в качестве вакцины ; более косвенно белок может управлять эндогенными стволовыми клетками, чтобы дифференцироваться желаемым образом.
Основные задачи РНК-терапии заключаются в доставке РНК в соответствующие клетки. Проблемы включают тот факт, что последовательности голой РНК естественным образом разрушаются после приготовления; они могут заставить иммунную систему организма атаковать их как захватчиков; и они непроницаемы для клеточной мембраны. Оказавшись внутри клетки, они должны покинуть транспортный механизм клетки, чтобы начать действовать в цитоплазме, в которой находятся необходимые рибосомы.
Преодолевая эти проблемы, мРНК в качестве терапевтического средства была впервые предложена в 1989 г. «после разработки широко применяемой техники трансфекции in vitro». В 1990-х годах были разработаны мРНК-вакцины от персонализированного рака, основанные на ненуклеозидной модифицированной мРНК. Терапия на основе мРНК продолжает изучаться как метод лечения или терапии как рака, так и аутоиммунных, метаболических и респираторных воспалительных заболеваний. Терапевтические методы редактирования генов, такие как CRISPR, также могут быть полезны при использовании мРНК для побуждения клеток к выработке желаемого белка Cas.
С 2010-х годов РНК-вакцины и другие РНК-терапевтические средства считаются «новым классом лекарств». Первые мРНК-вакцины на основе полученных ограниченного разрешения и были раскатывают по всему миру во время COVID-19 пандемии от Пфайзер-BioNTech COVID-19 вакцины и Moderna, например.
| У Схолии есть профиль информационной РНК (Q188928). |