
Войти
 Петля шпильки из пре-мРНК. Выделены азотистые основания (зеленый) и рибозофосфатный остов (синий). Это одиночная цепь РНК, которая сворачивается сама по себе.
Петля шпильки из пре-мРНК. Выделены азотистые основания (зеленый) и рибозофосфатный остов (синий). Это одиночная цепь РНК, которая сворачивается сама по себе. Рибонуклеиновая кислота (РНК ) представляет собой полимерную молекулу, которая играет важную роль в различных биологических ролях при кодировании , декодирование, регуляция и экспрессия генов . РНК и ДНК представляют собой нуклеиновые кислоты. Наряду с липидами, белками и углеводами нуклеиновые кислоты составляют одну из четырех основных макромолекул, необходимых для всех известных форм жизнь. Подобно ДНК, РНК собирается в виде цепочки из нуклеотидов, но в отличие от ДНК, РНК встречается в природе в виде одной цепи, свернутой на себя, а не в виде парной двойной цепи. Клеточные организмы используют информационную РНК (мРНК ) для передачи генетической информации (с использованием азотистых оснований из гуанина, урацила, аденин и цитозин, обозначаемые буквами G, U, A и C), которые направляют синтез определенных белков. Многие вирусы кодируют свою генетическую информацию с помощью РНК генома.
Некоторые молекулы РНК играют активную роль в клетках, катализируя биологические реакции, контролируя экспрессию гена или воспринимая и сообщая ответы на сотовые сигналы. Одним из этих активных процессов является синтез белка, универсальная функция, в которой молекулы РНК направляют синтез белков на рибосомах. В этом процессе используются молекулы транспортной РНК (тРНК ) для доставки аминокислот к рибосоме, где рибосомная РНК (рРНК ) затем соединяет аминокислоты вместе с образованием кодированных белков.
 Трехмерное представление рибосомной субъединицы 50S. Рибосомная РНК находится в охре, белки - в синем. Активный сайт представляет собой небольшой сегмент рРНК, обозначенный красным.
Трехмерное представление рибосомной субъединицы 50S. Рибосомная РНК находится в охре, белки - в синем. Активный сайт представляет собой небольшой сегмент рРНК, обозначенный красным. Химическая структура РНК очень похожа на структуру ДНК, но отличается по трем основным параметрам:
Подобно ДНК, большинство биологически активных РНК, включая мРНК, тРНК, рРНК, мяРНК и другие некодирующие РНК, содержат самокомплементарные последовательности, которые позволяют частям РНК складываться и соединяться между собой, образуя двойные спирали. Анализ этих РНК показал, что они высоко структурированы. В отличие от ДНК, их структуры не состоят из длинных двойных спиралей, а скорее представляют собой совокупность коротких спиралей, упакованных вместе в структуры, похожие на белки.
Таким образом, РНК могут достигать химического катализа (как ферменты). Например, определение структуры рибосомы - комплекса РНК-белок, который катализирует образование пептидной связи - показало, что ее активный сайт полностью состоит из РНК.
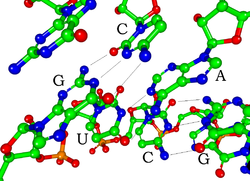 пар оснований Уотсона-Крика в миРНК (атомы водорода не показаны)
пар оснований Уотсона-Крика в миРНК (атомы водорода не показаны) Каждый нуклеотид в РНК содержит рибозный сахар с атомами углерода, пронумерованными от 1 'до 5'. Основание присоединено к положению 1 ', как правило, аденин (A), цитозин (C), гуанин (G) или урацил (U). Аденин и гуанин представляют собой пурины, цитозин и урацил представляют собой пиримидины. фосфатная группа присоединена к 3'-положению одной рибозы и 5'-положению следующей. Каждая фосфатная группа имеет отрицательный заряд, что делает РНК заряженной молекулой (полианионом). Основания образуют водородные связи между цитозином и гуанином, между аденином и урацилом и между гуанином и урацилом. Однако возможны и другие взаимодействия, такие как группа оснований аденина, связывающаяся друг с другом в выпуклости, или GNRA тетрапетля, которая имеет пару оснований гуанин-аденин.
 Структура фрагмента РНК, показывающая гуанозильную субъединицу.
Структура фрагмента РНК, показывающая гуанозильную субъединицу. Важным структурным компонентом РНК, который отличает ее от ДНК, является присутствие гидроксильной группы в положении 2 'рибозного сахара. Присутствие этой функциональной группы приводит к тому, что спираль в основном принимает геометрию А-формы, хотя в контексте одноцепочечных динуклеотидов РНК также редко может принимать B-форму, наиболее часто наблюдаемую в ДНК. Геометрия А-образной формы дает очень глубокую и узкую большую канавку и неглубокую и широкую малую канавку. Второе следствие присутствия 2'-гидроксильной группы состоит в том, что в конформационно гибких областях молекулы РНК (то есть, не участвующих в образовании двойной спирали) она может химически атаковать соседнюю фосфодиэфирную связь, чтобы расщепить основную цепь.
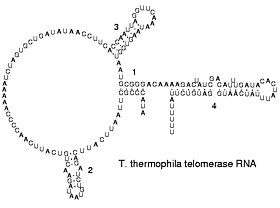 Вторичная структура теломеразной РНК.
Вторичная структура теломеразной РНК.РНК транскрибируется только с четырьмя основаниями (аденин, цитозин, гуанин и урацил), но эти основания и присоединенные сахара могут быть модифицированы многими способами, как РНК зрелый. Псевдоуридин (Ψ), в котором связь между урацилом и рибозой изменена с связи C – N на связь C – C, и риботимидин (T) встречаются в различных местах. (наиболее заметные из них находятся в петле TΨC тРНК ). Другое известное модифицированное основание - это гипоксантин, дезаминированное основание аденина, нуклеозид которого называется инозином (I). Инозин играет ключевую роль в гипотезе колебания генетического кода.
. Существует более 100 других природных модифицированных нуклеозидов. Наибольшее структурное разнообразие модификаций можно найти в тРНК, в то время как псевдоуридин и нуклеозиды с 2'-O-метилрибозой, часто присутствующие в рРНК, являются наиболее распространенными. Конкретные роли многих из этих модификаций в РНК до конца не изучены. Однако следует отметить, что в рибосомной РНК многие посттранскрипционные модификации происходят в высокофункциональных областях, таких как центр пептидилтрансферазы и интерфейс субъединицы, что означает, что они важны для нормального функционирования.
Функциональная форма одноцепочечных молекул РНК, как и белки, часто требует определенной третичной структуры. Каркас для этой структуры обеспечивается элементами вторичной структуры, которые представляют собой водородные связи внутри молекулы. Это приводит к нескольким распознаваемым «доменам» вторичной структуры, таким как петли шпильки, выпуклости и внутренние петли. Поскольку РНК заряжена, ионы металлов, такие как Mg, необходимы для стабилизации многих вторичных и третичных структур.
. Встречающийся в природе энантиомер РНК D -РНК, состоящая из D -рибонуклеотидов. Все центры хиральности расположены в D -рибозе. При использовании L -рибозы или, скорее, L -рибонуклеотидов, можно синтезировать L -РНК. L -РНК намного более устойчива против разложения РНКазой.
. Как и другие структурированные биополимеры, такие как белки, можно определить топологию свернутой молекулы РНК. Это часто делается на основе расположения внутрицепочечных контактов внутри свернутой РНК, что называется топологией цепи.
Синтез РНК обычно катализируется ферментом - РНК-полимеразой - использование ДНК в качестве матрицы, процесс, известный как транскрипция. Инициирование транскрипции начинается со связывания фермента с последовательностью промотора в ДНК (обычно находящейся «выше» гена). Двойная спираль ДНК разматывается за счет активности фермента геликазы. Затем фермент продвигается вдоль цепочки матрицы в направлении от 3 ’к 5’, синтезируя комплементарную молекулу РНК с удлинением в направлении от 5 ’к 3’. Последовательность ДНК также определяет, где будет происходить прекращение синтеза РНК.
Первичный транскрипт РНК часто модифицируются ферментами после транскрипции. Например, поли (А) хвост и 5'кэп добавляются к эукариотической пре-мРНК, а интроны удаляются сплайсосома.
Существует также ряд РНК-зависимых РНК-полимераз, которые используют РНК в качестве матрицы для синтеза новой цепи РНК. Например, ряд РНК-вирусов (таких как полиовирус) используют этот тип фермента для репликации своего генетического материала. Кроме того, РНК-зависимая РНК-полимераза является частью пути РНК-интерференции у многих организмов.
 Структура головки молотка рибозим, рибозим, разрезающий РНК.
Структура головки молотка рибозим, рибозим, разрезающий РНК. Информационная РНК (мРНК) - это РНК, несущая информацию от ДНК к рибосоме, участкам синтеза белка (трансляция ) в камере. Кодирующая последовательность мРНК определяет последовательность аминокислоты в продуцируемом белке. Однако многие РНК не кодируют белок (около 97% транскрипционной продукции у эукариот не кодирует белок).
Эти так называемые некодирующие РНК («нкРНК») могут кодироваться своими собственными генами (генами РНК), но также могут происходить из мРНК интронов. Наиболее яркими примерами некодирующих РНК являются транспортная РНК (тРНК) и рибосомная РНК (рРНК), обе из которых участвуют в процессе трансляции. Существуют также некодирующие РНК, участвующие в регуляции генов, процессинге РНК и другие роли. Некоторые РНК способны катализировать химические реакции, такие как разрезание и лигирование других молекул РНК, а также катализ образования пептидной связи в рибосоме ; они известны как рибозимы.
В соответствии с длиной цепи РНК, РНК включает малую РНК и длинную РНК. Обычно малые РНК короче 200 нуклеотидов в длину, а длинные РНК имеют длину более 200 нуклеотидов. Длинные РНК, также называемые большими РНК, в основном включают длинную некодирующую РНК (lncRNA) и мРНК. Малые РНК в основном включают 5.8S рибосомную РНК (рРНК), 5S рРНК, транспортную РНК (тРНК), микроРНК (миРНК), малая интерферирующая РНК (миРНК), малая ядрышковая РНК (мяРНК), взаимодействующая с Piwi РНК (пиРНК), малая РНК, полученная из тРНК (цРНК) и малая РНК, происходящая от рДНК (мрРНК). Есть определенные исключения, например, в случае 5S рРНК представителей рода Halococcus (Archaea ), которые имеют вставку, увеличивающую ее размер..
Информационная РНК (мРНК) несет информацию о последовательности белка в рибосомы, фабрики синтеза белка в клетке. Он имеет код , так что каждые три нуклеотида (кодон ) соответствуют одной аминокислоте. В эукариотических клетках, как только мРНК-предшественник (пре-мРНК) транскрибируется из ДНК, она процессируется до зрелой мРНК. Это удаляет его интроны - некодирующие участки пре-мРНК. Затем мРНК экспортируется из ядра в цитоплазму, где она связывается с рибосомами и транслируется в соответствующую белковую форму с помощью тРНК. В прокариотических клетках, которые не имеют компартментов ядра и цитоплазмы, мРНК может связываться с рибосомами, пока она транскрибируется с ДНК. Через некоторое время сообщение распадается на составляющие его нуклеотиды с помощью рибонуклеаз.
Передающая РНК (тРНК) представляет собой небольшую цепь РНК из примерно 80 нуклеотидов, которая передает специфическая аминокислота для растущей цепи полипептида в рибосомном сайте синтеза белка во время трансляции. Он имеет сайты для присоединения аминокислот и участок антикодона для распознавания кодона , который связывается с определенной последовательностью в цепи информационной РНК посредством водородных связей.
Рибосомная РНК ( рРНК) является каталитическим компонентом рибосом. Рибосомы эукариот содержат четыре различных молекулы рРНК: 18S, 5,8S, 28S и 5S рРНК. Три молекулы рРНК синтезируются в ядрышке, а одна синтезируется в другом месте. В цитоплазме рибосомная РНК и белок объединяются, образуя нуклеопротеин, называемый рибосомой. Рибосома связывает мРНК и осуществляет синтез белка. Несколько рибосом могут быть присоединены к одной мРНК в любое время. Почти вся РНК, обнаруженная в типичной эукариотической клетке, является рРНК.
РНК-мессенджер (тмРНК) обнаружена во многих бактериях и пластидах. Он маркирует белки, кодируемые мРНК, в которых отсутствуют стоп-кодоны для деградации, и предотвращает остановку рибосомы.
Самыми ранними известными регуляторами экспрессии гена были белки, известные как репрессоры и активаторы, регуляторы со специфическими короткими сайтами связывания в пределах энхансерных областей рядом с регулируемыми генами. Совсем недавно было обнаружено, что РНК также регулируют гены. У эукариот существует несколько видов РНК-зависимых процессов, регулирующих экспрессию генов в различных точках, таких как РНКи репрессирующие гены посттранскрипция союзник, длинные некодирующие РНК отключение блоков хроматина эпигенетически и энхансерных РНК, вызывающих повышенную экспрессию гена. В дополнение к этим механизмам у эукариот, как бактерии, так и археи, как было установлено, широко используют регуляторные РНК. Бактериальная малая РНК и система CRISPR являются примерами таких систем прокариотической регуляторной РНК. Файер и Мелло были удостоены Нобелевской премии по физиологии и медицине 2006 года за открытие микроРНК (миРНК), специфических коротких молекул РНК, которые могут образовывать пары оснований с мРНК.
Посттранскрипционные уровни экспрессии многих генов можно контролировать с помощью РНК-интерференции, в которой миРНК, специфические короткие молекулы РНК, соединяются с участками мРНК и нацелить их на деградацию. Этот основанный на антисмысловой процесс включает этапы, которые сначала обрабатывают РНК, чтобы она могла образовывать пару оснований с областью ее целевых мРНК. Как только происходит спаривание оснований, другие белки направляют мРНК на разрушение нуклеазами. За это открытие Файер и Мелло были удостоены Нобелевской премии по физиологии и медицине 2006 года.
Следующими за регуляцией стали Xist и другие длинные некодирующие РНК, связанные с инактивацией Х-хромосомы. Джинни Т. Ли и другие показали, что их роль, поначалу загадочная, заключалась в заглушении блоков хроматина посредством рекрутирования Polycomb комплекса, так что мессенджер С них нельзя было транскрибировать РНК. Было обнаружено, что дополнительные днРНК, в настоящее время определяемые как РНК из более чем 200 пар оснований, которые, по-видимому, не обладают кодирующим потенциалом, связаны с регуляцией стволовой клетки плюрипотентности и деления клеток..
Третья основная группа регуляторных РНК называется энхансерными РНК. В настоящее время неясно, являются ли они уникальной категорией РНК различной длины или представляют собой отдельное подмножество lncRNAs. В любом случае они транскрибируются с энхансеров, которые представляют собой известные регуляторные участки в ДНК рядом с генами, которые они регулируют. Они активируют транскрипцию гена (ов) под контролем энхансера, из которого они транскрибируются.
Сначала считалось, что регуляторная РНК является эукариотической Этот феномен является частью объяснения того, почему у высших организмов было обнаружено намного больше транскрипции, чем предполагалось. Но как только исследователи начали искать возможные регуляторы РНК в бактериях, они тоже обнаружили, что они называются малой РНК (мРНК). В настоящее время повсеместная природа систем регуляции РНК генов обсуждается в качестве подтверждения теории мира РНК. Бактериальные малые РНК обычно действуют посредством антисмыслового спаривания с мРНК, подавляющая ее трансляцию, либо влияя на стабильность, либо влияя на цис-связывающую способность. Были также обнаружены рибопереключатели. Они представляют собой цис-действующие регуляторные последовательности РНК, действующие аллостерически. Они меняют форму, когда связывают метаболиты, так что они приобретают или теряют способность связывать хроматин для регулирования экспрессии генов.
Археи также имеют системы регуляторных РНК. Система CRISPR, которая недавно использовалась для редактирования ДНК in situ, действует через регуляторные РНК у архей и бактерий, обеспечивая защиту от вирусных захватчиков.
 уридин в псевдоуридин является распространенной модификацией РНК. 359>Многие РНК участвуют в модификации других РНК. Интроны сплайсированы из пре-мРНК с помощью сплайсосом, которые содержат несколько малых ядерных РНК (мяРНК) или интроны могут быть рибозимами, которые сплайсируются сами по себе. РНК также может быть изменена путем модификации ее нуклеотидов до нуклеотидов, отличных от A, C, G и U. У эукариот модификации нуклеотидов РНК, как правило, направляются малыми ядрышковыми РНК (мяРНК; 60–300 нуклеотидов), обнаруженными в ядрышке и тельцах кахаля. snoRNA связываются с ферментами и направляют их к месту на РНК путем спаривания оснований с этой РНК. Затем эти ферменты выполняют модификацию нуклеотидов. рРНК и тРНК сильно модифицируются, но мяРНК и мРНК также могут быть целью модификации оснований. РНК также может быть метилирована. Hf РНК
уридин в псевдоуридин является распространенной модификацией РНК. 359>Многие РНК участвуют в модификации других РНК. Интроны сплайсированы из пре-мРНК с помощью сплайсосом, которые содержат несколько малых ядерных РНК (мяРНК) или интроны могут быть рибозимами, которые сплайсируются сами по себе. РНК также может быть изменена путем модификации ее нуклеотидов до нуклеотидов, отличных от A, C, G и U. У эукариот модификации нуклеотидов РНК, как правило, направляются малыми ядрышковыми РНК (мяРНК; 60–300 нуклеотидов), обнаруженными в ядрышке и тельцах кахаля. snoRNA связываются с ферментами и направляют их к месту на РНК путем спаривания оснований с этой РНК. Затем эти ферменты выполняют модификацию нуклеотидов. рРНК и тРНК сильно модифицируются, но мяРНК и мРНК также могут быть целью модификации оснований. РНК также может быть метилирована. Hf РНКПодобно ДНК, РНК может нести генетическую информацию. РНК-вирусы имеют геномы, состоящие из РНК, кодирующей ряд белков. Вирусный геном реплицируется некоторыми из этих белков, в то время как другие белки защищают геном, когда вирусная частица перемещается к новой клетке-хозяину. Вироиды - еще одна группа патогенов, но они состоят только из РНК, не кодируют какой-либо белок и реплицируются полимеразой клетки растения-хозяина.
Вирусы с обратной транскрипцией реплицируют свои геномы посредством обратной транскрипции копий ДНК из своей РНК; эти копии ДНК затем транскрибируются в новую РНК. Ретротранспозоны также распространяются путем копирования ДНК и РНК друг от друга, а теломераза содержит РНК, которая используется в качестве матрицы для построения концов хромосом эукариот.
 Двухцепочечная РНК
Двухцепочечная РНК Двухцепочечная РНК (дцРНК) - это РНК с двумя комплементарными цепями, подобная ДНК, обнаруженной во всех клетках, но с заменой тимина урацилом. дцРНК образует генетический материал некоторых вирусов (вирусов с двухцепочечной РНК ). Двухцепочечная РНК, такая как вирусная РНК или siRNA, может вызывать РНК-интерференцию у эукариот, а также реакцию интерферона у позвоночные животные.
В конце 1970-х было показано, что существует однониточная ковалентно замкнутая, то есть кольцевая форма РНК, экспрессируемая во всем царстве животных и растений (см. circRNA ). Считается, что circRNA возникают в результате реакции «обратного сплайсинга», когда сплайсосома присоединяется к нижележащему донору с расположенным выше акцепторным сайтом сплайсинга. До сих пор функция circRNAs в значительной степени неизвестна, хотя на нескольких примерах была продемонстрирована губчатая активность микроРНК.
 Роберт В. Холли (слева) со своей исследовательской группой.
Роберт В. Холли (слева) со своей исследовательской группой. Исследования РНК привели к множеству важных биологических открытий и множеству Нобелевских премий. Нуклеиновые кислоты были открыты в 1868 году Фридрихом Мишером, который назвал материал «нуклеином», поскольку он был обнаружен в ядре. Позже было обнаружено, что прокариотические клетки, не имеющие ядра, также содержат нуклеиновые кислоты. Роль РНК в синтезе белка подозревалась еще в 1939 году. Северо Очоа получил в 1959 Нобелевскую премию по медицине (совместно с Артуром Корнбергом ) после того, как открыл фермент, который может синтезировать РНК в лаборатории. Однако позднее было показано, что фермент, обнаруженный Очоа (полинуклеотидфосфорилаза ), отвечает за деградацию РНК, а не за синтез РНК. В 1956 году Алекс Рич и Дэвид Дэвис гибридизировали две отдельные цепи РНК, чтобы сформировать первый кристалл РНК, структура которого может быть определена с помощью рентгеновской кристаллографии.
Последовательность из 77 нуклеотидов тРНК дрожжей была обнаружена с помощью Роберт У. Холли в 1965 году, выиграв Холли Нобелевскую премию по медицине 1968 года (совместно с Хар Гобинд Хораной и Маршаллом Ниренбергом ).
В начале 1970-х годов были обнаружены ретровирусы и обратная транскриптаза, что впервые показало, что ферменты могут копировать РНК в ДНК (противоположный обычному пути для передача генетической информации). За эту работу Дэвид Балтимор, Ренато Дульбекко и Говард Темин были удостоены Нобелевской премии в 1975 году. В 1976 году Уолтер Фирс и его группа определила первую полную нуклеотидную последовательность генома РНК-вируса, последовательность бактериофага MS2.
. В 1977 году интроны и сплайсинг РНК были обнаружены как в вирусах млекопитающих, так и в в клеточных генах, в результате чего в 1993 г. получили Нобелевские премии Филип Шарп и Ричард Робертс. Каталитические молекулы РНК (рибозимы ) были открыты в начале 1980-х, что привело к присуждению Нобелевской премии 1989 года Томасу Чеху и Сидни Альтману. В 1990 году в Петунии было обнаружено, что введенные гены могут заглушать аналогичные гены самого растения, что, как теперь известно, является результатом РНК-интерференции.
Примерно в то же время длина 22 нт. Было обнаружено, что РНК, теперь называемые микроРНК, играют роль в развитии C. elegans. Исследования интерференции РНК были удостоены Нобелевской премии Эндрю Файра и Крейга Мелло в 2006 году, а еще одна Нобелевская премия была присуждена за исследования транскрипции РНК Роджеру Корнбергу в том же году. Открытие РНК, регулирующих гены, привело к попыткам разработать препараты на основе РНК, такие как siRNA, для подавления генов. В дополнение к Нобелевской премии, присужденной за исследования РНК в 2009 году, она была присуждена Венки Рамакришнану, Тому Стейтцу и Аде Йонат за выяснение атомной структуры рибосомы.
В 1968 году Карл Вёзе предположил, что РНК может быть каталитической, и предположил, что самые ранние формы жизни (самовоспроизводящиеся молекулы) могут полагались на РНК как для передачи генетической информации, так и для катализатора биохимических реакций - мир РНК.
В марте 2015 года комплекс ДНК и РНК нуклеотидов, включая урацил, цитозин и тимин, как сообщается, были образованы в лаборатории в условиях открытого космоса с использованием исходных химикатов, таких как пиримидин, органическое соединение, обычно обнаруживаемое в метеоритах. Пиримидин, как и полициклические ароматические углеводороды (ПАУ), является одним из наиболее богатых углеродом соединений, обнаруженных во Вселенной, и, возможно, был образован в красных гигантах или в межзвездной пыли и газовых облаках.
| В Wikiquote есть цитаты, относящиеся к: РНК |
| Викискладе есть средства массовой информации, связанные с РНК. |