
Войти
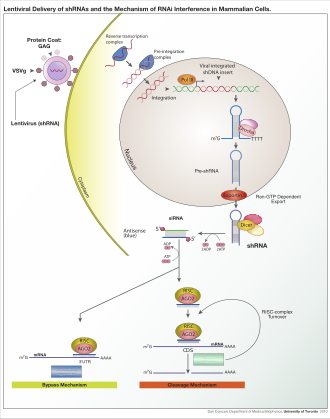 Лентивирусная доставка сконструированных кшРНК и механизм РНК-интерференции в клетках млекопитающих.
Лентивирусная доставка сконструированных кшРНК и механизм РНК-интерференции в клетках млекопитающих. РНК-интерференция (РНКи ) - это биологический процесс, в котором молекулы РНК подавляют экспрессию или трансляцию гена путем нейтрализации целевой мРНК молекул. Исторически РНКи были известны под другими названиями, включая совместное подавление, посттранскрипционное молчание генов (PTGS) и подавление. Детальное изучение каждого из этих, казалось бы, разных процессов выяснило, что идентичность этих явлений на самом деле была РНКи. Эндрю Файер и Крэйг С. Мелло разделили Нобелевскую премию по физиологии и медицине 2006 года за работу по вмешательству РНК в червя нематода Caenorhabditis elegans, который они опубликовали в 1998 году. С момента открытия РНКи и ее регуляторного потенциала стало очевидно, что РНКи обладает огромным потенциалом в подавлении желаемых генов. РНКи теперь известны как точные, эффективные, стабильные и более эффективные, чем антисмысловая терапия для подавления генов. Однако антисмысловая РНК, продуцируемая внутриклеточно с помощью вектора экспрессии, может быть разработана и найдет применение в качестве новых терапевтических агентов.
Два типа небольших молекул рибонуклеиновой кислоты (РНК) - микроРНК (miRNA) и малая интерферирующая РНК (siRNA ) - являются центральными для интерференции РНК. РНК являются прямыми продуктами генов, и эти малые РНК могут направлять ферментные комплексы на расщепление молекул матричной РНК (мРНК) и, таким образом, снижать их активность, предотвращая трансляцию, посредством посттранскрипционного сайленсинга генов. Более того, транскрипция может быть ингибирована с помощью механизма пре-транскрипционного сайленсинга РНК-интерференции, посредством которого ферментный комплекс катализирует метилирование ДНК в геномных положениях, комплементарных комплексной миРНК или миРНК. РНК-интерференция играет важную роль в защите клеток от паразитарных нуклеотидных последовательностей - вирусов и транспозонов. Он также влияет на развитие.
Путь РНКи обнаружен у многих эукариот, включая животных, и инициируется ферментом Dicer, который расщепляет длинные двойные разветвленная РНК (дцРНК) молекул в короткие двухцепочечные фрагменты ~ 21 нуклеотида миРНК. Каждая миРНК разматывается на две одноцепочечные РНК (оцРНК), пассажирскую цепь и направляющую цепь. Пассажирская цепь разрушается, а направляющая цепь включается в РНК-индуцированный комплекс сайленсинга (RISC). Наиболее хорошо изученным результатом является посттранскрипционное молчание гена, которое происходит, когда направляющая цепь соединяется с комплементарной последовательностью в молекуле матричной РНК и индуцирует расщепление аргонавтом 2 (Ago2), каталитическим компонентом RISC. У некоторых организмов этот процесс распространяется системно, несмотря на изначально ограниченные молярные концентрации миРНК.
РНКи является ценным инструментом исследования как в культуре клеток, так и в живых организмах, потому что синтетическая дцРНК, введенная в клетки, может селективно и надежно индуцировать супрессию конкретных представляющих интерес генов. РНКи можно использовать для крупномасштабных экранов, которые систематически отключают каждый ген в клетке, что может помочь идентифицировать компоненты, необходимые для конкретного клеточного процесса или события, такого как деление клетки. Этот путь также используется в качестве практического инструмента в биотехнологии, медицине и инсектицидах.
 Белок dicer из Giardia Кишечник, который катализирует расщепление дцРНК до миРНК. Домены РНКазы окрашены в зеленый цвет, домен PAZ - в желтый, платформенный домен - в красный, а соединительная спираль - в синий.
Белок dicer из Giardia Кишечник, который катализирует расщепление дцРНК до миРНК. Домены РНКазы окрашены в зеленый цвет, домен PAZ - в желтый, платформенный домен - в красный, а соединительная спираль - в синий. РНКи - это РНК-зависимый процесс подавления гена, который контролируется комплексом РНК-индуцированного сайленсинга (RISC) и инициируется короткими двухцепочечными молекулами РНК в цитоплазме клетки, где они взаимодействуют с каталитическим компонентом RISC аргонавтом. Когда дцРНК является экзогенной (возникшей в результате заражения вирусом с геномом РНК или лабораторных манипуляций), РНК импортируется непосредственно в цитоплазму и расщепляется на короткие фрагменты с помощью Dicer. Инициирующая дцРНК также может быть эндогенной (происходить из клетки), как в пре-микроРНК, экспрессируемых из генов, кодирующих РНК, в геноме. Первичные транскрипты таких генов сначала процессируются для образования характерной структуры стебель-петля пре-миРНК в ядре, а затем экспортируются в цитоплазму. Таким образом, два пути дцРНК, экзогенный и эндогенный, сходятся в RISC.
Экзогенная дцРНК инициирует РНКи путем активации рибонуклеазы белкового Дайсера, который связывает и расщепляет двухцепочечные РНК (дцРНК) в растениях или короткие шпилечные РНК (кшРНК) у людей для получения двухцепочечных фрагментов из 20–25 пар оснований с выступом из 2 нуклеотидов на 3'-конце. Биоинформатика исследования геномов множества организмов показывают, что такая длина максимизирует специфичность гена-мишени и сводит к минимуму неспецифические эффекты. Эти короткие двухцепочечные фрагменты называются малыми интерферирующими РНК (миРНК ). Эти миРНК затем разделяются на отдельные цепи и интегрируются в активный RISC с помощью комплекса загрузки RISC (RLC). RLC включает Dicer-2 и R2D2 и имеет решающее значение для объединения Ago2 и RISC. Фактор 11, связанный с TATA-связывающим белком (TAF11), собирает RLC, облегчая тетрамеризацию Dcr-2-R2D2, что увеличивает аффинность связывания с siRNA в 10 раз. Ассоциация с TAF11 преобразует комплекс R2-D2-инициатор (RDI) в RLC. R2D2 несет тандемные двухцепочечные РНК-связывающие домены для распознавания термодинамически стабильного конца дуплексов миРНК, тогда как Dicer-2 является другой менее стабильной конечностью. Загрузка асимметрична: домен MID Ago2 распознает термодинамически стабильный конец siRNA. Следовательно, «пассажирская» (смысловая) цепь, 5'-конец которой отбрасывается MID, выбрасывается, в то время как сохраненная «направляющая» (антисмысловая) цепь взаимодействует с AGO с образованием RISC.
После интеграции в RISC, миРНК соединяют пару оснований со своей целевой мРНК и расщепляют ее, тем самым предотвращая ее использование в качестве матрицы трансляции. В отличие от siRNA, RISC-комплекс, нагруженный miRNA, сканирует цитоплазматические мРНК на предмет потенциальной комплементарности. Вместо деструктивного расщепления (с помощью Ago2) miRNAs скорее нацелены на 3'-нетранслируемые области (UTR) мРНК, где они обычно связываются с несовершенной комплементарностью, тем самым блокируя доступ рибосом для трансляции.
Обнаруживается экзогенная дцРНК. и связывается эффекторным белком, известным как RDE-4 в C. elegans и R2D2 в Drosophila, которые стимулируют активность дайсера. Механизм, обеспечивающий такую специфичность длины, неизвестен, и этот белок связывает только длинные дцРНК.
У C. elegans этот инициирующий ответ усиливается посредством синтеза популяции «вторичных» миРНК, в течение которых инициирующие или «первичные» миРНК, продуцируемые дисером, используются в качестве матриц. Эти «вторичные» миРНК структурно отличаются от продуцируемых дайсером миРНК и, по-видимому, продуцируются РНК-зависимой РНК-полимеразой (RdRP).
 stem-loop вторичная структура пре- микроРНК из Brassica oleracea.
stem-loop вторичная структура пре- микроРНК из Brassica oleracea.MicroRNAs ( miRNA) - это геномно кодируемые некодирующие РНК, которые помогают регулировать экспрессию гена, особенно во время развития. Феномен РНК-интерференции, в широком смысле, включает в себя эндогенно индуцированные эффекты сайленсинга генов miРНК, а также сайленсинг, запускаемый чужеродной дцРНК. Зрелые миРНК структурно подобны миРНК, продуцируемым из экзогенной дцРНК, но до достижения зрелости миРНК должны сначала подвергнуться обширной посттранскрипционной модификации. MiRNA экспрессируется из гораздо более длинного гена, кодирующего РНК, в виде первичного транскрипта, известного как pri-miRNA, который процессируется в клеточном ядре до 70-нуклеотида структура стебель-петля, называемая пре-миРНК микропроцессорным комплексом . Этот комплекс состоит из фермента РНКазы III, называемого Drosha, и дцРНК-связывающего белка DGCR8. Часть dsRNA этой пре-miRNA связывается и расщепляется Dicer с образованием зрелой молекулы miRNA, которая может быть интегрирована в комплекс RISC; таким образом, миРНК и миРНК имеют один и тот же последующий клеточный аппарат. Во-первых, в EBV была описана кодируемая вирусом миРНК. После этого у вирусов было описано все большее количество микроРНК. VIRmiRNA - это полный каталог вирусных микроРНК, их мишеней и антивирусных miRNA (см. Также ресурс VIRmiRNA: http://crdd.osdd.net/servers/virmirna/).
миРНК, полученные из длинные предшественники дцРНК отличаются от miRNA тем, что miRNA, особенно у животных, обычно имеют неполное спаривание оснований с мишенью и ингибируют трансляцию многих различных мРНК со сходными последовательностями. Напротив, миРНК обычно идеально образуют пары оснований и индуцируют расщепление мРНК только в одной конкретной мишени. В Drosophila и C. elegans miРНК и siRNA процессируются различными белками аргонавта и дайсерами.
Три первичных нетранслируемых области (3'UTR) информационных РНК (мРНК) часто содержат регуляторные последовательности, которые посттранскрипционно вызывают интерференцию РНК. Такие 3'-UTR часто содержат как сайты связывания для микроРНК (miRNA), так и для регуляторных белков. Связываясь со специфическими сайтами в 3'-UTR, miRNA могут снижать экспрессию генов различных мРНК, либо ингибируя трансляцию, либо напрямую вызывая деградацию транскрипта. 3'-UTR также может иметь области сайленсеров, которые связывают репрессорные белки, которые ингибируют экспрессию мРНК.
3'-UTR часто содержит элементы ответа микроРНК (MRE). MRE - это последовательности, с которыми связываются miRNA. Это преобладающие мотивы в 3'-UTR. Среди всех регуляторных мотивов в 3'-UTR (например, включая области сайленсеров) MRE составляют около половины мотивов.
По состоянию на 2014 г. веб-сайт miRBase, архив последовательностей miRNA последовательностей и аннотаций, перечислил 28 645 записей о 233 биологических видах. Из них 1881 miRNA находились в аннотированных локусах miRNA человека. Предполагалось, что миРНК содержат в среднем около четырехсот целевых мРНК (влияющих на экспрессию нескольких сотен генов). Friedman et al. подсчитали, что>45000 сайтов-мишеней miRNA в 3'UTR мРНК человека консервативны выше фоновых уровней, и более 60% генов, кодирующих белок человека, находились под селективным давлением для поддержания спаривания с miRNA.
Прямые эксперименты показывают, что одна миРНК может снизить стабильность сотен уникальных мРНК. Другие эксперименты показывают, что одна миРНК может подавлять выработку сотен белков, но эта репрессия часто бывает относительно мягкой (менее чем в 2 раза).
Эффекты нарушения регуляции экспрессии генов miРНК кажутся важными. при раке. Например, при раке желудочно-кишечного тракта девять miRNA были идентифицированы как эпигенетически измененные и эффективные в подавлении регуляции ферментов репарации ДНК.
Эффекты дисрегуляции miRNA экспрессии генов также, по-видимому, важны для психоневрологические расстройства, такие как шизофрения, биполярное расстройство, большая депрессия, болезнь Паркинсона, болезнь Альцгеймера и расстройства аутистического спектра.
Экзогенная дцРНК выявляется и связывается эффекторным белком, известный как RDE-4 в C. elegans и R2D2 в Drosophila, которые стимулируют активность дайсера. Этот белок связывает только длинные дцРНК, но механизм, обеспечивающий специфичность этой длины, неизвестен. Этот РНК-связывающий белок затем облегчает перенос расщепленных миРНК в комплекс RISC.
У C. elegans этот ответ инициации усиливается за счет синтеза популяции «вторичных» миРНК, во время которых продуцируемые дайсером инициирующие или «первичные» миРНК используются в качестве матриц. Эти «вторичные» миРНК структурно отличаются от продуцируемых дайсером миРНК и, по-видимому, продуцируются РНК-зависимой РНК-полимеразой (RdRP).
 малая РНК Биогенез: первичные миРНК (при-миРНК) транскрибируются в ядре и складываются обратно в себя в виде шпилек, которые затем обрезаются в ядре микропроцессорным комплексом с образованием ~ 60-70nt шпилька пре-РНК. Эта пре-миРНК транспортируется через комплекс ядерных пор (NPC) в цитоплазму, где Дайсер дополнительно урезает ее до дуплекса миРНК ~ 20nt (пре-миРНК также входят в этот путь на этот шаг). Затем этот дуплекс загружается в Ago для образования «пре-RISC (комплекс подавления, индуцированного РНК)», и цепь-пассажир высвобождается с образованием активного RISC.
малая РНК Биогенез: первичные миРНК (при-миРНК) транскрибируются в ядре и складываются обратно в себя в виде шпилек, которые затем обрезаются в ядре микропроцессорным комплексом с образованием ~ 60-70nt шпилька пре-РНК. Эта пре-миРНК транспортируется через комплекс ядерных пор (NPC) в цитоплазму, где Дайсер дополнительно урезает ее до дуплекса миРНК ~ 20nt (пре-миРНК также входят в этот путь на этот шаг). Затем этот дуплекс загружается в Ago для образования «пре-RISC (комплекс подавления, индуцированного РНК)», и цепь-пассажир высвобождается с образованием активного RISC. Слева: полноразмерный аргонавтомат белок из архей вида Pyrococcus furiosus. Справа: домен PIWI белка аргонавта в комплексе с двухцепочечной РНК.
Слева: полноразмерный аргонавтомат белок из архей вида Pyrococcus furiosus. Справа: домен PIWI белка аргонавта в комплексе с двухцепочечной РНК.Активные компоненты РНК-индуцированного комплекса сайленсинга (RISC) эндонуклеазы, называемые белками аргонавтов, которые расщепляют цепь мРНК-мишени , комплементарную связанной с ними миРНК. Поскольку фрагменты, продуцируемые дайсером, являются двухцепочечными, каждый из них теоретически может продуцировать функциональную миРНК. Однако только одна из двух цепей, известная как направляющая цепь, связывает белок аргонавта и направляет подавление гена. Другая антинаправляющая нить или пассажирская нить разрушается во время активации RISC. Хотя сначала предполагалось, что АТФ -зависимая геликаза разделяет эти две цепи, оказалось, что этот процесс является АТФ-независимым и осуществляется непосредственно белковыми компонентами RISC. Однако кинетический анализ РНКи in vitro в присутствии и в отсутствие АТФ показал, что АТФ может потребоваться для раскручивания и удаления расщепленной цепи мРНК из комплекса RISC после катализа. Направляющая цепь, как правило, является той, у которой 5'-конец менее стабильно спарен с ее комплементом, но на выбор цепи не влияет направление, в котором дайсер расщепляет дцРНК до включения RISC. Вместо этого белок R2D2 может служить фактором дифференциации путем связывания более стабильного 5'-конца цепи-пассажира.
Структурная основа связывания РНК с белком аргонавта была исследована с помощью X- лучевая кристаллография связывающего домена РНК-связанного аргонавидного белка. Здесь фосфорилированный 5 'конец цепи РНК входит в консервативную основную поверхность карман и устанавливает контакты через двухвалентный катион (атом с двумя положительными зарядами), такой как магний, и ароматическим укладкой (процесс, который позволяет более чем один атом, чтобы поделиться электроном, передавая его вперед и назад) между 5'-нуклеотидом в миРНК и консервативным тирозиновым остатком. Считается, что этот сайт формирует сайт зародышеобразования для связывания миРНК с ее мРНК-мишенью. Анализ ингибирующего действия несовпадений на 5 'или 3' конце направляющей цепи показал, что 5'-конец направляющей цепи, вероятно, отвечает за сопоставление и связывание целевой мРНК, в то время как 3'-конец отвечает за физическое размещение мРНК-мишени в области RISC, благоприятной для расщепления.
Непонятно, как активированный комплекс RISC локализует комплементарные мРНК внутри клетки. Хотя было предложено, что процесс расщепления связан с трансляцией, трансляция мРНК-мишени не является существенной для опосредованной РНКи деградации. Действительно, РНКи могут быть более эффективными против мишеней мРНК, которые не транслируются. Белки Argonaute локализуются в определенных областях цитоплазмы, называемых P-тельцами (также цитоплазматических тельцах или GW-тельцах), которые представляют собой области с высокими скоростями распада мРНК; Активность miRNA также сгруппирована в P-тельцах. Нарушение Р-телец снижает эффективность РНК-интерференции, что позволяет предположить, что они являются критическим сайтом в процессе РНКи.
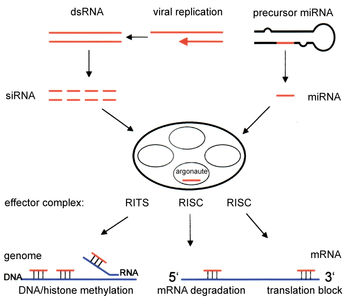 Фермент дайсер урезает двухцепочечную РНК, чтобы образуют малую интерферирующую РНК или микроРНК. Эти процессированные РНК включаются в РНК-индуцированный комплекс сайленсинга (RISC), который нацелен на информационную РНК для предотвращения трансляции.
Фермент дайсер урезает двухцепочечную РНК, чтобы образуют малую интерферирующую РНК или микроРНК. Эти процессированные РНК включаются в РНК-индуцированный комплекс сайленсинга (RISC), который нацелен на информационную РНК для предотвращения трансляции.Компоненты пути РНКи используются во многих эукариот в поддержании организации и структуры их геномов. Модификация гистонов и ассоциированная индукция образования гетерохроматина служит для подавления генов пре- транскрипционно ; этот процесс называется РНК-индуцированное подавление транскрипции (RITS) и осуществляется комплексом белков, называемым комплексом RITS. В делящихся дрожжах этот комплекс содержит аргонавт, белок Chp1 хромодомена и белок Tas3 неизвестной функции. Как следствие, для индукции и распространения гетерохроматиновых областей необходимы белки argonaute и RdRP. Действительно, делеция этих генов у делящихся дрожжей S. pombe нарушает метилирование гистонов и образование центромеры, вызывая медленную или остановленную анафазу во время деления клеток. В некоторых случаях наблюдались аналогичные процессы, связанные с модификацией гистонов, для активации транскрипции генов.
Механизм, с помощью которого комплекс RITS индуцирует образование и организацию гетерохроматина, недостаточно изучен. Большинство исследований было сосредоточено на области типа спаривания у делящихся дрожжей, которая не может быть репрезентативной для активности в других геномных областях / организмах. Поддерживая существующие участки гетерохроматина, RITS образует комплекс с сиРНК , комплементарным локальным генам, и стабильно связывает локальные метилированные гистоны, действуя котранскрипционно, разрушая любые возникающие транскрипты пре-мРНК, которые инициируются РНК-полимеразой. Формирование такой области гетерохроматина, хотя и не ее поддержание, зависит от дайсера, предположительно потому, что дайсер необходим для генерации начального комплемента миРНК, нацеленных на последующие транскрипты. Было высказано предположение, что поддержание гетерохроматина функционирует как самоусиливающаяся петля обратной связи, поскольку новые siRNAs формируются из случайных возникающих транскриптов с помощью RdRP для включения в локальные комплексы RITS. Актуальность наблюдений в областях и центромер спаривающегося типа делящихся дрожжей для млекопитающих неясна, поскольку поддержание гетерохроматина в клетках млекопитающих может не зависеть от компонентов пути РНКи.
Тип, который наиболее распространен у высших эукариот, превращает аденозин нуклеотиды в инозин в дцРНК с помощью фермента аденозиндезаминазы (ADAR). Первоначально в 2000 г. было высказано предположение, что пути редактирования РНКи и A → I РНК могут конкурировать за общий субстрат дцРНК. Некоторые пре-miRNAs подвергаются редактированию A → I РНК, и этот механизм может регулировать процессинг и экспрессию зрелых miRNA. Кроме того, по меньшей мере один ADAR млекопитающего может секвестировать миРНК из компонентов пути РНКи. Дальнейшее подтверждение этой модели исходит из исследований на ADAR-нулевых штаммах C. elegans, показывающих, что редактирование РНК A → I может противодействовать РНКи-сайленсингу эндогенных генов и трансгенов.
 Иллюстрация основных различий между молчанием генов растений и животных. Нативно экспрессируемая микроРНК или экзогенная малая интерферирующая РНК процессируется dicer и интегрируется в комплекс RISC, который опосредует подавление гена.
Иллюстрация основных различий между молчанием генов растений и животных. Нативно экспрессируемая микроРНК или экзогенная малая интерферирующая РНК процессируется dicer и интегрируется в комплекс RISC, который опосредует подавление гена. Организмы различаются по своей способности захватывать чужеродную дцРНК и использовать ее в пути РНКи. Эффекты РНК-интерференции могут быть как системными, так и наследственными у растений и C. elegans, но не у дрозофилы или млекопитающих. Считается, что в растениях РНКи размножаются путем передачи миРНК между клетками через плазмодесмы (каналы в клеточных стенках, которые обеспечивают связь и транспорт). Наследственность обусловлена метилированием промоторов, на которые нацелены РНКи; новый паттерн метилирования копируется в каждом новом поколении клетки. Общее различие между растениями и животными заключается в нацеливании на эндогенно продуцируемые miRNAs; у растений miRNAs обычно полностью или почти идеально комплементарны своим генам-мишеням и индуцируют прямое расщепление мРНК с помощью RISC, тогда как miRNA животных имеют тенденцию быть более дивергентными по последовательности и индуцируют репрессию трансляции. Этот эффект трансляции может быть вызван ингибированием взаимодействий факторов инициации трансляции с полиадениновым хвостом информационной РНК.
некоторыми эукариотическими простейшими, такими как Leishmania major и Trypanosoma cruzi полностью лишены пути РНКи. Большинство или все компоненты также отсутствуют у некоторых грибов, особенно у модельного организма Saccharomyces cerevisiae. Присутствие РНКи у других видов почкующихся дрожжей, таких как и Candida albicans, дополнительно демонстрирует, что индукция двух РНКи-родственных белков из S. castellii способствует РНКи в S. cerevisiae. Отсутствие у некоторых аскомицетов и базидиомицетов путей интерференции РНК указывает на то, что белки, необходимые для подавления РНК, были потеряны независимо от многих грибковых линий, возможно, из-за эволюции новый путь со схожей функцией или отсутствием избирательного преимущества в определенных нишах.
На экспрессию генов в прокариотах влияет система на основе РНК, в некоторых отношениях сходная с РНКи. Здесь гены, кодирующие РНК, контролируют изобилие или трансляцию мРНК, производя комплементарную РНК, которая отжигается с мРНК. Однако эти регуляторные РНК обычно не считаются аналогами миРНК, потому что в этом не участвует фермент дайсера. Было высказано предположение, что системы CRISPR-интерференции у прокариот аналогичны системам эукариотической РНК-интерференции, хотя ни один из белковых компонентов не ортологичен.
РНК-интерференция является жизненно важной частью иммунного ответа на вирусы и другой чужеродный генетический материал, особенно в растениях, где он также может предотвращать самораспространение транспозонов. Такие растения, как Arabidopsis thaliana экспрессируют несколько дайсеров гомологов, которые специализируются на различных реакциях, когда растение подвергается воздействию различных вирусов. Еще до того, как был полностью понят путь РНКи, было известно, что индуцированное молчание генов в растениях может распространяться по всему растению с системным эффектом и может передаваться из стада в растения привоя посредством прививки. Этот феномен с тех пор был признан особенностью адаптивной иммунной системы растений и позволяет всему растению реагировать на вирус после первоначального локализованного контакта. В ответ многие вирусы растений развили сложные механизмы для подавления ответа РНКи. К ним относятся вирусные белки, которые связывают короткие двухцепочечные фрагменты РНК с одноцепочечными выступающими концами, такие как те, которые продуцируются дайсером. Некоторые геномы растений также экспрессируют эндогенные миРНК в ответ на инфицирование конкретными типами бактерий. Эти эффекты могут быть частью общей реакции на патогены, которая подавляет любой метаболический процесс в организме хозяина, который способствует процессу инфекции.
Хотя животные обычно экспрессируют меньше вариантов фермента дайсера, чем растения, РНКи у некоторых животных продуцирует противовирусный ответ. Как у молодых, так и у взрослых дрозофил РНК-интерференция важна для противовирусного врожденного иммунитета и активна против патогенов, таких как вирус Drosophila X. Аналогичная роль в иммунитете может действовать и у C. elegans, поскольку белки аргонаута активируются в ответ на вирусы, а черви, избыточно экспрессирующие компоненты пути РНКи, устойчивы к вирусной инфекции.
Роль РНК-интерференции во врожденных процессах у млекопитающих. иммунитет плохо изучен, и имеется относительно мало данных. Однако существование вирусов, кодирующих гены, способные подавлять РНКи-ответ в клетках млекопитающих, может свидетельствовать в пользу РНКи-зависимого иммунного ответа млекопитающих, хотя эта гипотеза была оспорена как плохо обоснованная. Представлены доказательства существования функционального пути противовирусной РНКи в клетках млекопитающих.
Существуют также другие функции РНКи в вирусах млекопитающих, такие как миРНК, экспрессируемые вирусом герпеса, которые могут действовать поскольку гетерохроматин организация запускает вирусную латентность.
эндогенно экспрессируемых миРНК, включая как интронные, так и межгенные миРНК, наиболее важны для репрессии трансляции и регуляции развития, особенно в отношении сроков морфогенеза и поддержания недифференцированных или неполностью дифференцированных типов клеток, таких как стволовые ячейки. Роль эндогенно экспрессируемой миРНК в подавлении экспрессии гена была впервые описана у C. elegans в 1993 году. У растений эта функция была обнаружена, когда было показано, что «микроРНК JAW» Arabidopsis участвует в регуляции нескольких генов, контролирующих форму растений. В растениях большинство генов, регулируемых miRNA, являются факторами транскрипции ; таким образом, активность miRNA является особенно широкой и регулирует целые генные сети во время развития путем модуляции экспрессии ключевых регуляторных генов, включая факторы транскрипции, а также белки F-бокса. У многих организмов, включая человека, miRNA связаны с образованием опухолей и нарушением регуляции клеточного цикла. Здесь miRNA могут функционировать как онкогены и супрессоры опухолей.
На основании парсимонии филогенетического анализа, самый последний общий предок всех эукариот, скорее всего, уже обладал ранним путем РНК-интерференции; отсутствие этого пути у некоторых эукариот считается производной характеристикой. Эта предковая система РНКи, вероятно, содержала по крайней мере один дайсер-подобный белок, один аргонавт, один белок PIWI и РНК-зависимую РНК-полимеразу, которая также могла играть другие клеточные роли. Крупномасштабное исследование сравнительной геномики аналогичным образом показывает, что эукариотическая коронная группа уже обладала этими компонентами, которые, возможно, затем имели более тесные функциональные связи с общими системами деградации РНК, такими как экзосома. Это исследование также предполагает, что семейство РНК-связывающих белков аргонавтов, которое является общим для эукариот, большинства архей и, по крайней мере, некоторых бактерий (таких как Aquifex aeolicus ), гомологично и первоначально возникло из компонентов система инициирования перевода.
Обычно считается, что предковой функцией системы РНКи была иммунная защита от экзогенных генетических элементов, таких как транспозоны и вирусные геномы. Родственные функции, такие как модификация гистонов, возможно, уже присутствовали у предков современных эукариот, хотя другие функции, такие как регуляция развития с помощью miRNA, как полагают, развились позже.
Гены интерференции РНК как компоненты противовирусного Врожденная иммунная система у многих эукариот участвует в эволюционной гонке вооружений с вирусными генами. Некоторые вирусы развили механизмы для подавления ответа РНКи в своих клетках-хозяевах, особенно для вирусов растений. Исследования темпов эволюции у дрозофилы показали, что гены пути РНКи подвержены сильному направленному отбору и являются одними из самых быстро развивающихся генов в геноме дрозофилы.
Путь интерференции РНК часто используется в экспериментальной биологии для изучения функции генов в культуре клеток и in vivo в модельных организмах. Двухцепочечная РНК синтезируется с последовательностью, комплементарной интересующему гену, и вводится в клетку или организм, где она распознается как экзогенный генетический материал и активирует путь РНКи. Используя этот механизм, исследователи могут вызвать резкое снижение экспрессии целевого гена. Изучение эффектов этого снижения может показать физиологическую роль продукта гена. Поскольку РНКи не может полностью устранить экспрессию гена, этот метод иногда называют «нокдауном », чтобы отличить его от процедур «нокаут », в которых экспрессия гена полностью устранено. В недавнем исследовании проверка эффективности подавления РНКи с использованием данных массива генов показала 18,5% неудач в 429 независимых экспериментах.
Большие усилия в вычислительной биологии были направлены на разработку эффективных реагентов дцРНК. которые максимизируют нокдаун гена, но минимизируют «нецелевые» эффекты. Эффекты вне мишени возникают, когда введенная РНК имеет базовую последовательность, которая может спариваться и, таким образом, снижать экспрессию нескольких генов. Такие проблемы возникают чаще, когда дцРНК содержит повторяющиеся последовательности. Это было установлено на основе изучения геномов людей, C. elegans и S. pombe, что около 10% возможных миРНК имеют существенные эффекты, не соответствующие цели. Было разработано множество программных инструментов, реализующих алгоритмы для разработки общих специфичных для млекопитающих и вирусов siRNA, которые автоматически проверяются на возможную перекрестную реактивность.
В зависимости от организма и экспериментальной системы экзогенная РНК может представлять собой длинную цепь, предназначенную для расщепления с помощью дайсера, или короткие РНК, предназначенные для использования в качестве субстратов миРНК. В большинстве клеток млекопитающих используются более короткие РНК, поскольку длинные двухцепочечные молекулы РНК индуцируют ответ интерферона млекопитающих, форму врожденного иммунитета, которая неспецифически реагирует на чужеродный генетический материал. Мышиные ооциты и клетки ранних мышиных эмбрионов лишены этой реакции на экзогенную дцРНК и поэтому являются общей модельной системой для изучения эффектов нокдауна гена млекопитающих. Также были разработаны специализированные лабораторные методы для улучшения применимости РНКи в системах млекопитающих путем исключения прямого введения миРНК, например, посредством стабильной трансфекции плазмидой <408.>кодирующей соответствующую последовательность, из которой миРНК можно транскрибировать, или более сложными лентивирусными векторными системами, позволяющими индуцибельную активацию или дезактивацию транскрипции, известную как условная РНКи.
 Нормальная взрослая муха дрозофила, обычный модельный организм, используемый в экспериментах с РНКи.
Нормальная взрослая муха дрозофила, обычный модельный организм, используемый в экспериментах с РНКи. В большинстве применений РНКи в функциональной геномике у животных использовались C. elegans и Drosophila, поскольку это обычные модельные организмы, в которых РНКи наиболее эффективны. C. elegans особенно полезен для исследования РНКи по двум причинам: во-первых, эффекты сайленсинга генов обычно наследуются, а во-вторых, потому что доставка дцРНК чрезвычайно проста. Благодаря механизму, детали которого плохо изучены, бактерии, такие как E. coli, которые несут желаемую дцРНК, могут быть скармлены червям и будут передавать полезную нагрузку своей РНК червю через кишечник. Эта «доставка путем кормления» так же эффективна для индукции сайленсинга генов, как и более дорогие и трудоемкие методы доставки, такие как замачивание червей в растворе дцРНК и инъекция дцРНК в гонады. Хотя доставка является более сложной для большинства других организмов, также предпринимаются усилия по проведению крупномасштабного геномного скрининга в культуре клеток с клетками млекопитающих.
Подходы к созданию полногеномных библиотек РНКи могут потребовать более сложных, чем дизайн единственной миРНК для определенного набора экспериментальных условий. Искусственные нейронные сети часто используются для создания библиотек siRNA и для прогнозирования их вероятной эффективности при нокдауне гена. Массовый геномный скрининг широко рассматривается как многообещающий метод аннотации генома и инициировал разработку высокопроизводительных методов скрининга, основанных на микрочипах. Однако полезность этих скринингов и способность методов, разработанных на модельных организмах, распространяться даже на близкородственные виды, были поставлены под сомнение, например от C. elegans до родственных паразитических нематод.
Функциональная геномика с использованием РНКи является особенно привлекательный метод для геномного картирования и аннотации растений, поскольку многие растения являются полиплоидными, что представляет собой существенные проблемы для более традиционных методов генной инженерии. Например, РНКи успешно использовалась для исследований функциональной геномики мягкой пшеницы (которая является гексаплоидом), а также в более распространенных модельных системах растений Arabidopsis и кукурузы.
История использования РНКи в медицине
 Хронология использования РНКи в медицине между 1996 и 2017 годами
Хронология использования РНКи в медицине между 1996 и 2017 годами Первый случай РНК сайленсинга у животных был задокументирован в 1996 году, когда Гуо и Кемфуэс заметил, что, вводя смысл и антисмысловая РНК к мРНК par-1 в Caenorhabditis elegans вызывала деградацию сообщения par-1. Считалось, что эта деградация запускается одноцепочечной РНК (оцРНК), но два года спустя, в 1998 году, Файер и Мелло обнаружили, что эта способность подавлять экспрессию гена par-1 фактически запускается двухцепочечной РНК (дцРНК).). В конечном итоге они разделили Нобелевскую премию по физиологии и медицине за это открытие. Сразу после новаторского открытия Файра и Мелло Эльбашир и др. было обнаружено, что с помощью синтетически созданной малой интерферирующей РНК (siRNA) стало возможно нацеливаться на подавление определенных последовательностей в гене, а не на подавление всего гена. Только год спустя McCaffrey и его коллеги продемонстрировали, что это специфичное для последовательности молчание имеет терапевтическое применение, нацеливая последовательность из вируса гепатита C у трансгенных мышей. С тех пор несколько исследователей пытались расширить терапевтическое применение РНКи, в частности, пытаясь воздействовать на гены, вызывающие различные типы рака. К 2006 году первые приложения, прошедшие клинические испытания, были для лечения дегенерации желтого пятна и респираторно-синцитиального вируса. Четыре года спустя было начато первое клиническое испытание фазы I на людях с использованием системы доставки наночастиц для нацеливания на солидные опухоли. Хотя в настоящее время в большинстве исследований изучается применение РНКи в лечении рака, список возможных применений обширен. РНКи потенциально могут быть использованы для лечения вирусов, бактериальных заболеваний, паразитов, дезадаптивных генетических мутаций, контролировать потребление наркотиков, обезболивают и даже модулируют сон.
Противовирусное лечение - одно из первых предложенных медицинских приложений на основе РНКи, и были разработаны два разных типа. Первый тип направлен на вирусные РНК. Многие исследования показали, что нацеливание вирусных РНК может подавлять репликацию множества вирусов, включая ВИЧ, ВПЧ, гепатит A, гепатит B, вирус гриппа, респираторно-синцитиальный вирус (RSV), коронавирус SARS (SARS-CoV), аденовирус и вирус кори. Другая стратегия состоит в том, чтобы заблокировать начальные проникновения вируса путем нацеливания на гены клетки-хозяина. Например, подавление хемокиновых рецепторов (CXCR4 и CCR5 ) в клетках-хозяевах может предотвратить проникновение вируса ВИЧ.
В то время как традиционный химиотерапия может эффективно убивать раковые клетки, отсутствие специфичности для различения нормальных клеток и раковых клеток при таком лечении обычно вызывает серьезные побочные эффекты. Многочисленные исследования продемонстрировали, что РНКи может обеспечить более специфический подход к подавлению роста опухоли, воздействуя на гены, связанные с раком (т.е. онкоген ). Также было высказано предположение, что РНКи может повышать чувствительность раковых клеток к химиотерапевтическим агентам, обеспечивая комбинаторный терапевтический подход с химиотерапией. Другим потенциальным лечением на основе РНКи является ингибирование клеточной инвазии и миграции.
Стратегии РНКи также показывают потенциал для лечения нейродегенеративных заболеваний. Исследования на клетках и мышах показали, что специфическое воздействие РНКи на гены, продуцирующие амилоид бета (например, BACE1 и APP), может значительно снизить количество пептида Aβ, которое коррелирует с причиной болезни Альцгеймера.. Кроме того, эти подходы, основанные на подавлении звука, также обеспечивают многообещающие результаты в лечении болезни Паркинсона и болезни полиглутамина.
Для достижения клинического потенциала RNAi, миРНК должна эффективно транспортироваться к клеткам тканей-мишеней. Однако существуют различные препятствия, которые необходимо устранить, прежде чем его можно будет использовать в клинических условиях. Например, «голая» миРНК восприимчива к нескольким препятствиям, которые снижают ее терапевтическую эффективность. Кроме того, как только миРНК попадает в кровоток, голая РНК может расщепляться нуклеазами сыворотки и стимулировать врожденную иммунную систему. Из-за своего размера и сильно полианионной (содержащей отрицательные заряды на нескольких сайтах) природы немодифицированные молекулы миРНК не могут легко проникать в клетки через клеточную мембрану. Следовательно, необходимо использовать искусственные или инкапсулированные в наночастицы миРНК. Однако транспортировка siRNA через клеточную мембрану по-прежнему имеет свои уникальные проблемы. Если миРНК переносится через клеточную мембрану, может возникнуть непреднамеренная токсичность, если терапевтические дозы не оптимизированы, и миРНК могут проявлять эффекты, не соответствующие цели (например, непреднамеренное подавление генов с частичной комплементарностью последовательностей ). Даже после попадания в клетки требуется повторное дозирование, поскольку их эффекты ослабляются при каждом делении клеток. Как описано ранее, части вектора, транспортирующие дцРНК, также могут иметь регуляторные эффекты. Таким образом, следует учитывать и контролировать неспецифические побочные эффекты.
По сравнению с химиотерапией или другими противораковыми препаратами, препарат siRNA имеет много преимуществ. SiRNA действует на посттранскрипционной стадии экспрессии генов, поэтому она не модифицирует и не изменяет ДНК, оказывая вредное воздействие. SiRNA также можно использовать для получения определенного ответа определенным способом, например, путем снижения уровня подавления экспрессии генов. В одной раковой клетке siRNA может вызывать резкое подавление экспрессии генов всего с несколькими копиями. Это происходит путем подавления генов, способствующих развитию рака, с помощью РНКи, а также нацеливания на последовательность мРНК.
Лекарства с РНКи лечат рак, подавляя определенные гены, способствующие развитию рака. Это достигается путем дополнения раковых генов РНКи, например, сохранения последовательностей мРНК в соответствии с препаратом РНКи. В идеале РНКи следует вводить и / или химически модифицировать, чтобы РНКи могли более эффективно достигать раковых клеток. Поглощение и регуляция РНКи контролируется почками.
Иммунная система человека делится на две отдельные ветви: врожденная иммунная система и адаптивная иммунная система. Врожденная иммунная система является первой защитой от инфекции и реагирует на патогены обычным образом. С другой стороны, адаптивная иммунная система, система, которая была развита позже, чем врожденная, состоит в основном из высокоспециализированных В- и Т-клеток, которые обучены реагировать на определенные части патогенных молекул.
Проблема между старыми патогенами и новыми помогла создать систему защищенных клеток и частиц, которые называются безопасным каркасом. Эта структура дала людям армейские системы, которые ищут и уничтожают частицы-захватчики, такие как патогены, микроскопические организмы, паразиты и инфекции. Безопасный каркас для млекопитающих был разработан для включения миРНК в качестве инструмента для индикации вирусного заражения, что позволило миРНК создать интенсивный врожденный иммунный ответ.
миРНК контролируется врожденной иммунной системой, которую можно разделить на острые воспалительные реакции и противовирусные реакции. Воспалительный ответ создается с помощью сигналов от небольших сигнальных молекул или цитокинов. К ним относятся интерлейкин-1 (IL-1), интерлейкин-6 (IL-6), интерлейкин-12 (IL-12) и фактор некроза опухоли α (TNF-α). Врожденная иммунная система генерирует воспаление и противовирусные реакции, которые вызывают рецепторы распознавания паттернов высвобождения (PRR). Эти рецепторы помогают определить, какие патогены являются вирусами, грибами или бактериями. Более того, важность siRNA и врожденной иммунной системы состоит в том, чтобы включать больше PRR, чтобы помочь распознавать различные структуры РНК. Это увеличивает вероятность того, что siRNA вызовет иммуностимулирующий ответ в случае патогена.
Клинические исследования фаз I и II терапии siRNA, проведенные между 2015 и 2017 продемонстрировали мощный и устойчивый нокдаун гена в печени, с некоторыми признаками клинического улучшения и без неприемлемой токсичности. В настоящее время проводятся два исследования фазы III по лечению семейных нейродегенеративных и сердечных синдромов, вызванных мутациями в транстиретине (TTR). Многочисленные публикации показали, что системы доставки in vivo очень перспективны и разнообразны по характеристикам, что позволяет использовать их в самых разных областях. Система доставки наночастиц является наиболее многообещающей, однако этот метод создает дополнительные проблемы в производственном процессе, такие как необходимость жестко контролируемых процессов смешивания для достижения стабильного качества лекарственного препарата.
В таблице ниже показаны различные препараты, использующие РНК-интерференцию, и их фазы и статус в клинических испытаниях по состоянию на 2013 год.
| Лекарство | Цель | Система доставки | Заболевание | Фаза | Статус | Компания | Идентификатор |
| ALN – VSP02 | KSP и VEGF | LNP | Солидные опухоли | I | Завершенные | Alnylam Pharmaceuticals | NCT01158079 |
| siRNA – EphA2 – DOPC | EphA2 | LNP | Рак на поздних стадиях | I | Набор персонала | Онкологический центр MD Anderson | NCT01591356 |
| Atu027 | PKN3 | LNP | Солидные опухоли | I | Завершено | Silence Therapeutics | NCT00938574 |
| TKM –080301 | PLK1 | LNP | Рак | I | Набор персонала | Tekmira Pharmaceutical | NCT01262235 |
| TKM – 100201 | VP24, VP35, Заир L-полимераза Эбола | LNP | инфекция, вызванная вирусом Эбола | I | Вербовка | Tekmira Pharmaceutical | NCT01518881 |
| ALN – RSV01 | нуклеокапсид RSV | Naked siRNA | Респираторно-синцитиальный вирус инфекции | II | Завершено | Alnylam Pharmaceuticals | NCT00658086 |
| PRO-040201 | ApoB | LNP | Гиперхолестеринемия | I | Прекращена | Tekmira Pharmaceutical | NCT00927459 |
| ALN – PCS02 | PCSK9 | LNP | Гиперхолестеринемия | I | Завершено | Alnylam Pharmaceuticals | NCT01437059 |
| ALN – TTR02 | TTR | LNP | Транстиретин- опосредованный амилоидоз | II | Рекрутинг | Alnylam Pharmaceuticals | NCT01617967 |
| CALAA-01 | RRM2 | Циклодекстрин NP | Солидные опухоли | I | Активные | Calando Pharmaceuticals | NCT00689065 |
| TD101 | K6a (мутация N171K) | Голая миРНК | Pachyonychia congenita | I | Завершено | Pachyonychia Congenita Project | NCT00716014 |
| AGN211745 | VEGFR1 | Naked siRNA | Возрастная дегенерация желтого пятна, хориоидальная неоваскуляризация | II | Прекращено | Allergan | NCT00395057 |
| QPI-1007 | CASP2 | Голая siRNA | Атрофия зрительного нерва, неартериальная передняя ишемическая оптическая нейропатия | I | Завершено | Quark Pharmaceuticals | NCT01064505 |
| I5NP | p53 | Naked siRNA | Повреждение почек, острая почечная недостаточность | I | Завершено | Quark Pharmaceuticals | NCT00554359 |
| Задержка функции трансплантата, осложнения трансплантата почки | I, II | Рекрутинг | Quark Pharmaceuticals | NCT00802347 | |||
| PF-655 (PF-04523655) | RTP801 (проприетарный tar get) | Голая siRNA | Неоваскуляризация хориоидеи, диабетическая ретинопатия, диабетический макулярный отек | II | Active | Quark Pharmaceuticals | NCT01445899 |
| siG12D LODER | KRAS | полимер LODER | рак поджелудочной железы | II | Рекрутинг | Silenseed | NCT01676259 |
| Бевасираниб | VEGF | Голая миРНК | Диабетический отек желтого пятна, дегенерация желтого пятна | II | Завершено | Opko Health | NCT00306904 |
| SYL1001 | TRPV1 | Голая siRNA | Боль в глазах, сухость- глазной синдром | I, II | Рекрутинг | Sylentis | NCT01776658 |
| SYL040012 | ADRB2 | Naked siRNA | Глазная гипертензия, открытоугольная глаукома | II | Рекрутинг | Sylentis | NCT01739244 |
| CEQ508 | CTNNB1 | кшРНК, несущая Escherichia coli | Семейный аденоматозный полип osis | I, II | Рекрутинг | Marina Biotech | Неизвестно |
| RXi-109 | CTGF | Самодоставляющееся соединение РНКи | Профилактика рубцов | I | Рекрутинг | RXi Pharmaceuticals | NCT01780077 |
| ALN – TTRsc | TTR | конъюгат миРНК-GalNAc | Транстиретин-опосредованный амилоидоз | I | Рекрутинг | Alnylam Pharmaceuticals | NCT01814839 |
| ARC-520 | Консервированные области HBV | DPC | HBV | I | Рекрутинг | Arrowhead Research | NCT01872065 |
Было использовано вмешательство РНК для приложений в биотехнологии и приближается к коммерциализации в других областях. RNAi привела к изобретению новых сельскохозяйственных культур, таких как безникотиновый табак, кофе без кофеина, растения, обогащенные питательными веществами, и гипоаллергенные культуры. Генетически модифицированные арктические яблоки получили одобрение FDA в 2015 году. Яблоки были произведены путем подавления РНКи гена PPO (полифенолоксидазы), в результате чего получаются сорта яблок, которые не потемнеют после нарезки. Яблоки с замкнутым PPO не могут превращать хлорогеновую кислоту в стандартный хиноновый продукт.
Есть несколько возможностей для применения РНКи в растениеводстве для ее улучшения, таких как устойчивость к стрессу и повышение уровня питательности. RNAi докажет свой потенциал ингибирования фотодыхания для повышения продуктивности растений C3. Эта нокдаун-технология может быть полезна для стимулирования раннего цветения, отсроченного созревания, замедленного старения, нарушения покоя, снятия стресса растений, преодоления самостерильности и т. Д.
RNAi использовалась для генетически сконструировать растения для производства более низких уровней естественных токсинов растений. Такие методы используют стабильный и наследуемый фенотип РНКи в растительных стадах. Семена хлопка богаты диетическим белком, но, естественно, содержат токсичный терпеноид продукт госсипол, что делает их непригодными для потребления человеком. РНКи использовалась для производства хлопковых культур, семена которых содержат пониженные уровни дельта-кадинен-синтазы, ключевого фермента в производстве госсипола, без влияния на производство фермента в других частях растения, где госсипол сам по себе важен для предотвращение повреждения растений вредителями. Аналогичные усилия были направлены на уменьшение содержания цианогенного натурального продукта линамарина в растениях кассавы.
Нет растительных продуктов, в которых используется РНКи генная инженерия еще вышла из экспериментальной стадии. Усилия по разработке позволили успешно снизить уровни аллергенов в растениях томатов и обогатить такие растения, как томаты, диетическими антиоксидантами. Предыдущие коммерческие продукты, включая томат Flavr Savr и два сорта устойчивой к кольцевой пятнистости папайи, были первоначально разработаны с использованием антисмысловых технология, но, вероятно, использовала путь РНКи. РНКи сайленсинг альфа-амилазы также использовался для уменьшения роста грибов Aspergillus flavus кукурузы, которые в противном случае загрязнили бы ядра опасными афлатоксинами. Подавление синтазы слезоточивого фактора в луке привело к образованию лука без слез, и РНКи была использована в генах BP1 в рапсе для улучшения фотосинтеза. Гены SBEIIa и SBEIIb в пшенице были нацелены на пшеницу, чтобы производить более высокие уровни амилозы для улучшения функции кишечника.
Еще одна попытка снизила предшественники вероятных канцерогенов в растениях табака. Другие свойства растений, которые были разработаны в лаборатории, включают производство ненаркотических ненаркотических натуральных продуктов с помощью опийного мака и устойчивость к обычным вирусам растений.
РНКи находится в стадии разработки как инсектицид с использованием нескольких подходов, включая генную инженерию и местное применение. Клетки в средней кишке некоторых насекомых захватывают молекулы дцРНК в процессе, называемом РНКи окружающей среды. У некоторых насекомых эффект носит системный характер, поскольку сигнал распространяется по телу насекомого (называемый системной РНКи).
Животные, подвергшиеся воздействию РНКи в дозах, в миллионы раз превышающих ожидаемые уровни воздействия на человека, не проявляют никаких побочных эффектов.
РНКи по-разному действует на разные виды Lepidoptera (бабочки и мотыльки). Возможно, потому, что их слюна и сок кишечника лучше расщепляют РНК, хлопковая совка, совка-совка и азиатский рисовый мотыль до сих пор не доказана их чувствительность к РНКи при кормлении.
Недавние данные свидетельствуют о том, что устойчивость к РНКи может быть широкого спектра, что означает, что устойчивость к одной последовательности может придавать устойчивость к другим последовательностям дцРНК. В одной лабораторной популяции западного кукурузного корневого червя устойчивость возникла из-за недостаточного поглощения дцРНК DvSnf7 через кишечник. Когда другие последовательности дцРНК были протестированы против DvSnf7, другие последовательности перестали быть эффективными, что предполагает, что управление устойчивостью будет более трудным, чем простое отключение последовательностей дцРНК. Сочетание нескольких стратегий, таких как конструирование белка Cry, полученного из бактерии Bacillus thuringiensis (Bt), и РНКи в одном растении задерживают начало устойчивости.
Трансгенные культуры были созданы для экспрессии дцРНК, тщательно подобранной, чтобы заставить замолчать важнейшие гены у вредителей-мишеней. Эти дцРНК предназначены для воздействия только на насекомых, которые экспрессируют определенные генные последовательности. В качестве доказательства принципа, в 2009 году исследование показало, что РНК могут убить один из четырех видов плодовых мух, не нанося вреда трем другим.
В 2012 году Syngenta купила бельгийскую RNAi фирму Devgen за 522 миллиона долларов и Monsanto заплатила 29,2 миллиона долларов за исключительные права на интеллектуальную собственность у Alnylam Pharmaceuticals. Международный центр картофеля в Лиме, Перу занимается поиском генов, нацеленных на сладкого картофельного долгоносика, жука, личинки которого уничтожают сладкий картофель во всем мире. Другие исследователи пытаются заставить замолчать гены муравьев, гусениц и пыльцевых жуков. Monsanto, вероятно, будет первой на рынке с трансгенным семенем кукурузы, которое экспрессирует дцРНК на основе гена Snf7 западного кукурузного корневого червя, жука, личинки которого ежегодно вызывают ущерб в один миллиард долларов только в Соединенных Штатах. В статье 2012 года было показано, что подавление Snf7 останавливает рост личинок, убивая их в считанные дни. В 2013 году та же команда показала, что РНК влияет на очень немногие другие виды.
В качестве альтернативы дцРНК может поставляться без генной инженерии. Один из подходов - добавить их в поливную воду. Молекулы всасываются в сосудистую систему растений и отравляют насекомых, питающихся ими. Другой подход включает распыление дцРНК, как обычного пестицида. Это позволит быстрее адаптироваться к сопротивлению. Такие подходы потребуют недорогих источников дцРНК, которые в настоящее время не существуют.
Исследование РНКи на уровне генома основано на высокопроизводительном скрининге (HTS) технологии. Технология RNAi HTS позволяет проводить скрининг потери функции по всему геному и широко используется для идентификации генов, связанных с конкретными фенотипами. Эта технология была провозглашена потенциальной второй волной геномики после первой волны геномики платформ открытия экспрессии генов микроматрицы и однонуклеотидного полиморфизма. Одним из основных преимуществ скрининга РНКи на уровне генома является его способность одновременно опрашивать тысячи генов. Благодаря способности генерировать большой объем данных за эксперимент, скрининг РНКи в масштабе генома привел к взрывному росту скорости генерации данных. Использование таких больших наборов данных является фундаментальной проблемой, требующей подходящих методов статистики / биоинформатики. Основной процесс клеточного скрининга РНКи включает выбор библиотеки РНКи, надежных и стабильных типов клеток, трансфекцию РНКи агентами, обработку / инкубацию, обнаружение сигнала, анализ и идентификацию важных генов или терапевтических мишеней.
 Пример петуния растения, в которых гены пигментации подавляются РНКи. Левое растение - дикого типа ; правильные растения содержат трансгены, которые вызывают подавление экспрессии как трансгена, так и эндогенного гена, что приводит к появлению непигментированных белых участков цветка.
Пример петуния растения, в которых гены пигментации подавляются РНКи. Левое растение - дикого типа ; правильные растения содержат трансгены, которые вызывают подавление экспрессии как трансгена, так и эндогенного гена, что приводит к появлению непигментированных белых участков цветка. Процесс РНКи упоминается как «совместная супрессия» и «подавление», когда наблюдается до знания механизма, связанного с РНК. Открытию РНКи предшествовали, прежде всего, наблюдения за ингибированием транскрипции антисмысловой РНК, экспрессируемой в трансгенных растениях, и, более конкретно, сообщениями о неожиданных результатах в экспериментах, проведенных специалистами по растениеводству в США и Нидерланды в начале 1990-х годов. В попытке изменить цвет цветка в петунии, исследователи ввели дополнительные копии гена, кодирующего халконсинтазу, ключевой фермент пигментации цветов в растения петунии с обычно розовыми или фиолетовыми цветками. Ожидалось, что сверхэкспрессия гена приведет к появлению более темных цветов, но вместо этого у некоторых цветов будет менее заметный пурпурный пигмент, иногда в виде пестрого рисунка, что указывает на то, что активность халконсинтазы была существенно снижена или стала подавляться в зависимости от контекста. Позднее это будет объяснено как результат вставки трансгена рядом с промоторами в противоположном направлении в различных положениях по всему геному некоторых трансформантов, что приводит к экспрессии антисмысловых транскриптов и подавлению гена, когда эти промоторы активны. Еще одно раннее наблюдение РНКи было получено в ходе исследования грибка Neurospora crassa, хотя не сразу было признано, что это родство. Дальнейшее исследование феномена у растений показало, что подавление происходило из-за посттранскрипционного ингибирования экспрессии генов за счет увеличения скорости деградации мРНК. Это явление было названо совместным подавлением экспрессии генов, но молекулярный механизм остался неизвестным.
Вскоре после этого вирусологи растений, работающие над повышением устойчивости растений к вирусным заболеваниям, наблюдали похожее неожиданное явление. Хотя было известно, что растения, экспрессирующие вирус-специфические белки, демонстрируют повышенную толерантность или устойчивость к вирусной инфекции, не ожидалось, что растения, несущие только короткие, некодирующие области последовательностей вирусной РНК, будут демонстрировать аналогичные уровни защиты. Исследователи полагали, что вирусная РНК, продуцируемая трансгенами, также может подавлять репликацию вируса. Обратный эксперимент, в котором короткие последовательности генов растений были введены в вирусы, показал, что целевой ген подавлялся в инфицированном растении. Этот феномен был назван «индуцированным вирусом сайленсингом генов» (VIGS), а совокупность таких явлений получила общее название посттранскрипционного сайленсинга генов.
После этих первоначальных наблюдений на растениях лаборатории искали этот феномен у других организмов.. Крейг С. Мелло и Эндрю Файер в статье Nature 1998 года сообщили о мощном эффекте подавления генов после инъекции двухцепочечной РНК в C. elegans. При исследовании регуляции выработки мышечного белка они обнаружили, что ни инъекции мРНК, ни антисмысловой РНК не влияли на продукцию белка, но двухцепочечная РНК успешно подавляла глушитель целевого гена. В результате этой работы они придумали термин RNAi. Это открытие представляло собой первую идентификацию возбудителя этого явления. Файер и Мелло были удостоены Нобелевской премии по физиологии и медицине 2006 г. .
| На Wikimedia Commons есть материалы, связанные с РНК-интерференцией. |
| В Викиверситете есть учебные ресурсы по РНК-интерференции |