
Войти
 Структура E. coli геликаза RuvA
Структура E. coli геликаза RuvA | ДНК-геликаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 3.6.4.12 | ||||||||
| Базы данных | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| KEGG | Запись KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| PRIAM | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
| РНК-геликаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер EC | 3.6.4.13 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | Запись BRENDA | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| KEGG | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| PRIAM | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
геликазы - это класс ферментов, жизненно важных для всех организмов. Их основная функция - распаковывать гены организма. Это моторные белки, которые перемещаются направленно вдоль нуклеиновой кислоты фосфодиэфирного остова, разделяя две отожженные нити нуклеиновых кислот. такие как ДНК и РНК (следовательно, гелик- + -аза ), используя энергию от АТФ гидролиз. Существует множество геликаз, представляющих большое разнообразие процессов, в которых необходимо катализировать разделение цепей. Примерно 1% генов эукариот кодируют геликазы. геном человека кодирует 95 неизбыточных геликаз: 64 РНК-геликазы и 31 ДНК-геликазы. Многие клеточные процессы, такие как репликация ДНК, транскрипция, трансляция, рекомбинация, репарация ДНК и биогенез рибосом включает разделение цепей нуклеиновых кислот, что требует использования геликаз.
Геликазы часто используются для разделения цепей ДНК двойная спираль или самоотжигающаяся молекула РНК, использующая энергию гидролиза АТФ, процесса, характеризующегося расщеплением водорода связывает между отожженными нуклеотидными основаниями. Они также действуют для удаления белков, связанных с нуклеиновыми кислотами, и катализируют гомологичную рекомбинацию ДНК. Хеликазы способствуют метаболическим процессам РНК, таким как трансляция, транскрипция, биогенез рибосом, сплайсинг РНК, транспорт РНК и деградация РНК. Геликазы постепенно перемещаются вдоль одной цепи нуклеиновой кислоты дуплекса с направленностью и процессивностью, специфичными для каждого конкретного фермента.
Геликазы принимают разные структуры и состояния олигомеризации. В то время как DnaB -подобные геликазы раскручивают ДНК в виде кольцевых гексамеров, было показано, что другие ферменты активны в виде мономеров или димеры. Исследования показали, что геликазы могут действовать пассивно, ожидая некатализируемого раскручивания и затем перемещаясь между смещенными цепями, или могут играть активную роль в катализе разделения цепей с использованием энергии, генерируемой при гидролизе АТФ. В последнем случае геликаза действует аналогично активному двигателю, раскручиваясь и перемещаясь по субстрату как прямой результат ее АТФазной активности. Геликазы могут обрабатываться намного быстрее in vivo, чем in vitro из-за присутствия дополнительных белков, которые способствуют дестабилизации вилочного соединения.
 Действие геликазы при репликации ДНК
Действие геликазы при репликации ДНК Действие ферментной геликазы, например, раскручивающихся нуклеиновых кислот, достигается за счет снижения активационного барьера (





Факторы, которые влияют на высоту активационного барьера, включают: специфическую последовательность нуклеиновой кислоты вовлеченной молекулы, количество задействованных пар оснований, натяжение репликационной вилки и силы дестабилизации.
Размер активационного барьера, преодолеваемого геликазой, способствует ее классификации как активная или пассивная геликаза. В пассивных геликасах существует значительный барьер активации (определяемый как 




В активных геликазах,
Активные геликазы демонстрируют сходное поведение при воздействии на двухцепочечные нуклеиновые кислоты, дцНК или оцНК, в отношении скорости раскрутки и скорости транслокации, где в обеих системах
Эти две категории геликаз также можно моделировать как механизмы. В таких моделях пассивные геликазы концептуализируются как броуновские трещотки, управляемые тепловыми флуктуациями и последующими анизотропными градиентами по решетке ДНК. Активные геликасы, напротив, концептуализируются как шаговые двигатели - также известные как электродвигатели с силовым ходом - для продвижения используются либо конформационный «дюймовой червяк», либо ручной «шагающий» механизм. В зависимости от организма такое продвижение по спирали может происходить при скорости вращения в диапазоне от 5000 до 10000 об / мин.
ДНК-геликазы были обнаружены в E. coli в 1976 году. Эта геликаза была описана как «фермент, раскручивающий ДНК», который, как «обнаружено, денатурирует дуплексы ДНК в АТФ- зависимая реакция без заметного разложения ». Первая эукариотическая ДНК-геликаза была обнаружена в 1978 году в лилии. С тех пор ДНК-геликазы были обнаружены и выделены у других бактерий, вирусов, дрожжей, мух и высших эукариот. К настоящему времени выделено по меньшей мере 14 различных геликаз из одноклеточных организмов, 6 геликаз из бактериофагов, 12 из вирусов, 15 из дрожжей, 8 из растений, 11 из тимуса теленка и приблизительно 25 геликаз из клеток человека. Ниже приводится история открытия геликазы:
Общая функция геликаз за тот факт, что они демонстрируют определенную степень гомологии последовательности аминокислот ; все они имеют мотивы последовательности, расположенные внутри их первичной структуры, участвующие в связывании АТФ, гидролизе АТФ и транслокации вдоль нуклеиновая кислота субстрат. Вариабельная часть последовательности аминокислоты связана со специфическими особенностями каждой геликазы.
Присутствие этих мотивов геликазы позволяет отнести предполагаемую активность геликазы к данному белку, но не обязательно подтверждает ее как активную геликазу. Консервативные мотивы, однако, подтверждают эволюционную гомологию ферментов. На основе этих мотивов геликазы был выделен ряд суперсемейств геликаз.
Геликазы классифицируются на 6 групп (суперсемейства) на основе их общих мотивов последовательностей. Геликазы, не образующие кольцевой структуры, входят в суперсемейства 1 и 2, а образующие кольца геликазы входят в суперсемейства с 3 по 6. Геликазы также классифицируются как α или β в зависимости от того, работают ли они с одноцепочечной или двухцепочечной ДНК ; α-геликазы работают с одноцепочечной ДНК, а β-геликазы работают с двухцепочечной ДНК. Их также классифицируют по полярности транслокаций. Если происходит транслокация 3’-5 ’, геликаза относится к типу A; если происходит транслокация 5’-3 ’, это тип B.
Все геликазы являются членами P-петли или содержащего мотив Уокера семья.
Ген ATRX кодирует АТФ-зависимую геликазу, ATRX (также известную как XH2 и XNP) семейства подгруппы SNF2, которое, как полагают, отвечает за такие функции, как ремоделирование хроматина, регуляция генов и метилирование ДНК. Эти функции помогают предотвратить апоптоз, что приводит к регуляции размера коры, а также способствует выживанию гиппокампа и корковых структур, влияя на память и обучение. Эта геликаза расположена на Х-хромосоме (Xq13.1-q21.1) в перицентромерном гетерохроматине и связывается с белком гетерохроматина 1. Исследования показали, что ATRX играет роль в метилировании рДНК и важен для эмбрионального развития. Мутации были обнаружены во всем белке ATRX, причем более 90% из них расположены в доменах цинкового пальца и геликазы. Мутации ATRX могут приводить к X-связанной-альфа-талассемии - умственной отсталости (синдром ATR-X ).
Было обнаружено, что различные типы мутаций, обнаруженные в ATRX, связаны с ATR-X, в том числе наиболее часто одиночные - базовые миссенс-мутации, а также бессмысленные мутации, мутации сдвига рамки считывания и делеционные мутации. Характеристики ATR-X включают: микроцефалию, скелетные и лицевые аномалии, умственную отсталость, генитальные аномалии, судороги, ограниченное использование языка и способности и альфа-талассемию. Фенотип Наблюдаемое в ATR-X предполагает, что мутация гена ATRX вызывает подавление экспрессии генов, таких как гены альфа-глобина. До сих пор неизвестно, что вызывает экспрессию различных характеристик ATR-X у разных пациентов.
XPD (фактор D Xeroderma pigmentosum, также известный как белок ERCC2) представляет собой 5'-3 ', суперсемейство II, АТФ-зависимую геликазу, содержащую кластерные домены железо-сера. мутации в XPD геликазе hav Было показано, что он связан с нарушениями ускоренного старения, такими как синдром Кокейна (CS) и трихотиодистрофия (TTD). Синдром Кокейна и трихотиодистрофия являются нарушениями развития, включающими чувствительность к ультрафиолетовому излучению и преждевременное старение, а синдром Коккейна проявляет серьезную умственную отсталость с момента рождения. Мутация геликазы XPD также была причастна к xeroderma pigmentosum (XP), заболеванию, характеризующемуся чувствительностью к УФ-свету и приводящему к увеличению в несколько тысяч раз развития рака кожи.
XPD является важным компонентом комплекса TFIIH, фактора транскрипции и репарации в клетке. В составе этого комплекса он способствует эксцизионной репарации нуклеотидов путем раскручивания ДНК. TFIIH помогает восстанавливать поврежденную ДНК, например поврежденную солнцем. Мутация геликазы XPD, которая помогает формировать этот комплекс и способствует его функции, вызывает чувствительность к солнечному свету, наблюдаемую при всех трех заболеваниях, а также повышенный риск рака, наблюдаемый при XP, и преждевременное старение, наблюдаемое при трихотиодистрофии и синдроме Кокейна.
Мутации геликазы XPD, приводящие к трихотиодистрофии, обнаруживаются по всему белку в различных местах, участвующих в белок-белковых взаимодействиях. Эта мутация приводит к нестабильному белку из-за его неспособности образовывать стабилизирующие взаимодействия с другими белками в точках мутаций. Это, в свою очередь, дестабилизирует весь комплекс TFIIH, что приводит к дефектам механизмов транскрипции и репарации клетки.
Было высказано предположение, что мутации геликазы XPD, приводящие к синдрому Кокейна, могут быть результатом мутаций внутри XPD., вызывая жесткость белка и последующую неспособность переключиться с функций репарации на функции транскрипции из-за «блокировки» в режиме репарации. Это могло заставить геликазу разрезать сегменты ДНК, предназначенные для транскрипции. Хотя текущие данные указывают на дефект геликазы XPD, приводящий к потере гибкости белка в случаях синдрома Кокейна, все еще неясно, как эта структура белка приводит к симптомам, описанным при синдроме Кокейна.
В xeroderma pigmentosa, мутация геликазы XPD существует в месте связывания АТФ или ДНК. Это приводит к структурно-функциональной геликазе, способной облегчить транскрипцию, однако она подавляет ее функцию по раскручиванию ДНК и репарации ДНК. Отсутствие способности клетки восстанавливать мутации, например, вызванные солнечным повреждением, является причиной высокого уровня заболеваемости раком у пациентов с пигментной ксеродермией.
 геликаза RecQ
геликаза RecQ геликаза RecQ (3'-5 ') относятся к группе геликаз суперсемейства II, которые помогают поддерживать стабильность генома и подавляют несоответствующую рекомбинацию. Дефициты и / или мутации в геликазах семейства RecQ демонстрируют аберрантную генетическую рекомбинацию и / или репликацию ДНК, что приводит к хромосомной нестабильности и общему снижению способности к пролиферации. Было показано, что мутации в геликазах семейства RecQ BLM, RECQL4 и WRN, которые играют роль в регуляции гомологичной рекомбинации, приводят к аутосомно-рецессивным заболеваниям синдром Блума (BS), синдром Ротмунда-Томсона (RTS) и синдром Вернера (WS), соответственно.
Синдром Блума характеризуется предрасположенностью к раку с ранним началом, со средним возраст начала заболевания 24 года. У пациентов с синдромом Блума наблюдается высокая частота реципрокного обмена между сестринскими хроматидами (SCE) и чрезмерное повреждение хромосом. Имеются данные, позволяющие предположить, что BLM играет роль в восстановлении нарушенной репликации ДНК в ответвлениях репликации.
Синдром Вернера - это нарушение преждевременного старения с симптомами, включая раннее начало атеросклероза и остеопороза и других возрастных заболеваний, a высокая частота саркомы и смерть, часто наступающая от инфаркта миокарда или рака на 4-6-м десятилетии жизни. Клетки пациентов с синдромом Вернера демонстрируют сокращенную продолжительность репродуктивной жизни из-за хромосомных разрывов и транслокаций, а также большие делеции хромосомных компонентов, вызывающие нестабильность генома.
синдром Ротмунда-Томсона, также известный как врожденная пойкилодермия, характеризуется преждевременным старением, кожными и скелетными аномалиями, сыпью, пойкилодермой, ювенильной катарактой и предрасположенностью к онкологическим заболеваниям, таким как остеосаркомы. Хромосомные перестройки, вызывающие нестабильность генома, обнаружены в клетках пациентов с синдромом Ротмунда-Томсона.
Во время мейоза двухцепочечные разрывы ДНК и другие ДНК Повреждения в хроматиде устраняются посредством гомологичной рекомбинации с использованием либо сестринской хроматиды, либо гомологичной несестринской хроматиды в качестве матрицы. Эта репарация может привести к кроссоверу (CO) или, что более часто, к рекомбинанту без кроссовера (NCO). В дрожжах Schizosaccharomyces pombe ДНК-геликаза FmI1 семейства FANCM направляет образование рекомбинации NCO во время мейоза. Хеликаза RecQ-типа Rqh1 также направляет мейотическую рекомбинацию NCO. Эти геликазы, благодаря своей способности раскручивать промежуточные соединения D-петли, способствуют рекомбинации NCO посредством процесса зависимого от синтеза отжига цепи.
В растении Arabidopsis thaliana, FANCM геликаза способствует NCO и противодействует образованию рекомбинантов CO. Другая геликаза, RECQ4A / B, также независимо снижает CO. Было высказано предположение, что CO ограничены из-за долгосрочных затрат на рекомбинацию CO, то есть разрушение благоприятных генетических комбинаций аллелей, созданных прошлым естественным отбором.
 Human DEAD-box РНК-геликаза
Human DEAD-box РНК-геликаза 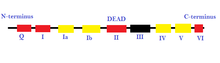 На этом изображении представлены различные промоторные последовательности и дополнительные домены, которые способствуют раскручиванию РНК (локальное разделение цепей). Области, выделенные красным, представляют собой домены связывания АТФ, а области, отмеченные желтым цветом, - домены взаимодействия РНК. Присутствуют также специфические последовательности, называемые DEAD-бокс-белками, которые помогают катализировать реакции, в которых нет необходимости в прямом гидролизе АТФ, если он связывается с доменами цепи.
На этом изображении представлены различные промоторные последовательности и дополнительные домены, которые способствуют раскручиванию РНК (локальное разделение цепей). Области, выделенные красным, представляют собой домены связывания АТФ, а области, отмеченные желтым цветом, - домены взаимодействия РНК. Присутствуют также специфические последовательности, называемые DEAD-бокс-белками, которые помогают катализировать реакции, в которых нет необходимости в прямом гидролизе АТФ, если он связывается с доменами цепи. РНК-геликазы необходимы для большинства процессов метаболизма РНК, например как биогенез рибосомы, сплайсинг пре-мРНК и инициация трансляции. Они также играют важную роль в обнаружении вирусных РНК. РНК-геликазы участвуют в обеспечении противовирусного иммунного ответа, поскольку они могут идентифицировать чужеродные РНК у позвоночных. Около 80% всех вирусов являются РНК-вирусами и содержат собственные РНК-геликазы. Дефектные РНК-геликазы связывают с раком, инфекционными заболеваниями и нейродегенеративными расстройствами. Некоторые неврологические расстройства, связанные с дефектными РНК-геликазами: боковой амиотрофический склероз, спинальная мышечная атрофия, спиноцеребеллярная атаксия 2 типа, болезнь Альцгеймера и синдром летальной врожденной контрактуры..
РНК-геликазы и ДНК-геликазы могут быть обнаружены вместе во всех суперсемействах геликаз, кроме SF6. Все эукариотические РНК-геликазы, которые были идентифицированы на сегодняшний день, не образуют кольца и являются частью SF1 и SF2. С другой стороны, кольцеобразные РНК-геликазы были обнаружены у бактерий и вирусов. Однако не все РНК-геликазы проявляют геликазную активность, определяемую ферментативной функцией, т.е. белки семейства Swi / Snf. Хотя эти белки несут типичные мотивы геликазы, гидролизуют АТФ зависимым от нуклеиновых кислот образом и построены вокруг ядра геликазы, в целом, никакой раскручивающей активности не наблюдается.
РНК-геликазы, которые действительно проявляют раскручивающую активность, обладают характеризуется как минимум двумя различными механизмами: каноническим разматыванием дуплекса и локальным разделением цепей. Каноническая дуплексная раскрутка - это ступенчатое направленное разделение дуплексной цепи, как описано выше, для раскручивания ДНК. Однако локальное разделение цепей происходит в процессе, при котором фермент геликаза загружается в любом месте дуплекса. Обычно этому способствует одноцепочечный участок РНК, а загрузка фермента сопровождается связыванием АТФ. Как только геликаза и АТФ связываются, происходит локальное разделение цепи, которое требует связывания АТФ, но не фактического процесса гидролиза АТФ. Дуплекс, представленный меньшим количеством пар оснований, затем диссоциирует без дополнительной помощи со стороны фермента. Этот режим раскрутки используется DEAD / DEAH-бокс-геликазами.
. В настоящее время в Интернете доступна база данных РНК-геликаз, которая содержит полный список РНК-геликаз с такой информацией, как последовательность, структура, биохимические и клеточные функции.
Для измерения активности геликазы in vitro используются различные методы. Эти методы варьируются от качественных (анализы, которые обычно приводят к результатам, не включающим значений или измерений) до количественных (анализы с числовыми результатами, которые можно использовать в статистическом и числовом анализе). В 1982–1983 годах был разработан первый прямой биохимический анализ для измерения активности геликазы. Этот метод получил название «анализ смещения нити».
Другие методы были позже применены. разработан, который включает в себя некоторые, если не все из следующего: высокопроизводительную механику, использование нерадиоактивной маркировки нуклеотидов, более быстрое время реакции / меньшее потребление времени, мониторинг активности геликазы в реальном времени (с использованием кинетического измерения вместо конечной точки / одиночной точечный анализ). Эти методологии включают: «метод быстрого гашения потока, анализы на основе флуоресценции, анализы фильтрации, сцинтилляционный анализ близости, анализ с временным разрешением флуоресцентный резонансный перенос энергии, анализ на основе флэш-планшетов. технологии, гомогенные анализы гашения флуоресценции с временным разрешением и анализы геликазы на основе электрохемилюминесценции ". С использованием специализированных математических уравнений некоторые из этих анализов можно использовать для определения, сколько спаренных оснований нуклеотидов хеликаза может разрушить за гидролиз 1 молекулы АТФ.
Также доступны коммерчески доступные диагностические наборы. Одним из таких наборов является диагностический анализ Trupoint от PerkinElmer, Inc. Этот анализ представляет собой анализ тушения флуоресценции с временным разрешением, в котором используется технология PerkinEmer «SignalClimb», основанная на двух метках, которые связываются в непосредственной близости. друг к другу, но на противоположных цепях ДНК. Одна метка представляет собой флуоресцентный хелат лантаноида, который служит меткой, за которой следят с помощью адекватного 96/384-луночного ридера для планшетов. Другая метка - это молекула органического тушителя. В основе этого анализа лежит «гашение» или подавление сигнала хелата лантаноида органической молекулой гасителя, когда они находятся в непосредственной близости - как это было бы, когда дуплекс ДНК находится в нативном состоянии. При активности геликазы на дуплексе метки гасителя и лантанида разделяются по мере разматывания ДНК. Эта потеря близости сводит на нет способность гасителей подавлять сигнал лантаноида, вызывая обнаруживаемое увеличение флуоресценции, которое является репрезентативным для количества размотанной ДНК и может использоваться в качестве количественного измерения активности геликазы. Выполнение и использование методов флуоресцентной визуализации одиночных молекул с упором на методы, которые включают оптический захват в сочетании с эпифлуоресцентной визуализацией, а также иммобилизацию поверхности в сочетании с визуализацией флуоресценции полного внутреннего отражения. В сочетании с микроканальными проточными кюветами и микрожидкостным контролем позволяет визуализировать и отслеживать отдельные флуоресцентно меченые молекулы белка и ДНК, обеспечивая измерение раскручивания и транслокации ДНК с разрешением по одной молекуле.
Полярность геликазы, которую также называют «направленностью», определяется как направление (характеризуемое как 5 '→ 3' или 3 '→ 5') движения геликазы на однонитевой ДНК / РНК, вдоль которой она движется. Это определение полярности жизненно важно, например, определение, прикрепляется ли тестируемая геликаза к ведущей цепи ДНК или к отстающей цепи ДНК. Чтобы охарактеризовать эту особенность геликазы, частично дуплексная ДНК используется в качестве субстрата, который имеет центральную одноцепочечную область ДНК с различной длиной дуплексных областей ДНК (одна короткая область, которая проходит 5 '→ 3', и одна более длинная область, которая проходит через 3 ''). '→ 5') по обе стороны от этой области. Как только геликаза добавляется к этой центральной одноцепочечной области, полярность определяется характеристикой вновь образованной одноцепочечной ДНК.
| Викискладе есть медиафайлы, связанные с Helicases. |