
Войти
 В мейозе хромосома или хромосомы дублируются (во время интерфазы ), а гомологичные хромосомы обмениваются генетической информацией ( хромосомный кроссовер ) во время первого деления, называемого мейозом I. Дочерние клетки снова делятся в мейозе II, разделяя сестринские хроматиды с образованием гаплоидных гамет. Две гаметы сливаются во время оплодотворения, образуя диплоидную клетку с полным набором парных хромосом.
В мейозе хромосома или хромосомы дублируются (во время интерфазы ), а гомологичные хромосомы обмениваются генетической информацией ( хромосомный кроссовер ) во время первого деления, называемого мейозом I. Дочерние клетки снова делятся в мейозе II, разделяя сестринские хроматиды с образованием гаплоидных гамет. Две гаметы сливаются во время оплодотворения, образуя диплоидную клетку с полным набором парных хромосом.  Воспроизвести медиа Видео мейоза I в сперматоцитах мухи журавля, воспроизведенное со скоростью 120 × записанной
Воспроизвести медиа Видео мейоза I в сперматоцитах мухи журавля, воспроизведенное со скоростью 120 × записанной Мейоз ( / м aɪ oʊ s ɪ s / ( слушать ) ![]() , от греческого μείωσις, мейоз, что означает «ослабление», потому что это восстановительное деление) представляет собой особый тип клеточного деления из зародышевых клеток в сексуально-размножающихся организмах используются для производят гаметы, такие как сперматозоиды или яйцеклетки. Он включает в себя два раунда деления, в результате которых образуются четыре клетки с одной копией каждой хромосомы ( гаплоидом ). Кроме того, перед делением генетический материал от отцовской и материнской копий каждой хромосомы скрещивается, создавая новые комбинации кода на каждой хромосоме. Позже, во время оплодотворения, гаплоидные клетки, произведенные мейозом у мужчины и женщины, сливаются, чтобы снова создать клетку с двумя копиями каждой хромосомы, зиготу.
, от греческого μείωσις, мейоз, что означает «ослабление», потому что это восстановительное деление) представляет собой особый тип клеточного деления из зародышевых клеток в сексуально-размножающихся организмах используются для производят гаметы, такие как сперматозоиды или яйцеклетки. Он включает в себя два раунда деления, в результате которых образуются четыре клетки с одной копией каждой хромосомы ( гаплоидом ). Кроме того, перед делением генетический материал от отцовской и материнской копий каждой хромосомы скрещивается, создавая новые комбинации кода на каждой хромосоме. Позже, во время оплодотворения, гаплоидные клетки, произведенные мейозом у мужчины и женщины, сливаются, чтобы снова создать клетку с двумя копиями каждой хромосомы, зиготу.
Ошибки в мейозе, приводящие к анеуплоидии (аномальному количеству хромосом), являются ведущей известной причиной выкидыша и наиболее частой генетической причиной нарушений развития.
В мейозе репликация ДНК сопровождается двумя раундами клеточного деления с образованием четырех дочерних клеток, каждая из которых имеет половину хромосом по сравнению с исходной родительской клеткой. Два подразделения мейоза известны как мейоз I и мейоз II. Перед мейоз начинается во время S фазы от клеточного цикла, ДНК каждой хромосомы реплицируется, так что она состоит из двух идентичных сестринских хроматид, которые по- прежнему удерживаются вместе с помощью когезии сестринских хроматид. Эта S-фаза может упоминаться как «премейотическая S-фаза» или «мейотическая S-фаза». Сразу после репликации ДНК мейотические клетки вступают в длительную G 2- подобную стадию, известную как мейотическая профаза. В это время гомологичные хромосомы соединяются друг с другом и подвергаются генетической рекомбинации - запрограммированному процессу, в котором ДНК может быть разрезана, а затем восстановлена, что позволяет им обмениваться некоторой генетической информацией. Подмножество событий рекомбинации приводит к кроссоверам, которые создают физические связи, известные как хиазмы (единственное число: хиазма, от греческой буквы Хи (X) ) между гомологичными хромосомами. У большинства организмов эти связи могут помочь каждой паре гомологичных хромосом отделиться друг от друга во время Мейоза I, в результате чего образуются две гаплоидные клетки, которые имеют половину количества хромосом по сравнению с родительской клеткой.
Во время мейоза II сцепление между сестринскими хроматидами ослабляется, и они отделяются друг от друга, как во время митоза. В некоторых случаях все четыре мейотических продукта образуют гаметы, такие как сперма, споры или пыльца. У самок животных три из четырех продуктов мейоза обычно удаляются путем экструзии в полярные тельца, и только одна клетка развивается, чтобы произвести яйцеклетку. Поскольку количество хромосом уменьшается вдвое во время мейоза, гаметы могут сливаться (то есть оплодотворяться ), образуя диплоидную зиготу, которая содержит две копии каждой хромосомы, по одной от каждого родителя. Таким образом, чередующиеся циклы мейоза и оплодотворения обеспечивают половое размножение, при этом последующие поколения поддерживают одинаковое количество хромосом. Например, диплоидные клетки человека содержат 23 пары хромосом, включая 1 пару половых хромосом (всего 46), половину материнского происхождения и половину отцовского происхождения. Мейоз производит гаплоидные гаметы (яйцеклетки или сперматозоиды), которые содержат один набор из 23 хромосом. Когда две гаметы (яйцеклетка и сперматозоид) сливаются, образующаяся зигота снова становится диплоидной, причем каждая мать и отец вносят по 23 хромосомы. Такой же паттерн, но не одинаковое количество хромосом, встречается у всех организмов, использующих мейоз.
Мейоз встречается у всех одноклеточных и многоклеточных организмов, размножающихся половым путем (все они являются эукариотами ), включая животных, растения и грибы. Это важный процесс для оогенеза и сперматогенеза.
Хотя процесс мейоза связан с более общим процессом деления клеток митоза, он отличается в двух важных отношениях:
| рекомбинация | мейоз | перетасовывает гены между двумя хромосомами в каждой паре (по одному от каждого родителя), создавая рекомбинантные хромосомы с уникальными генетическими комбинациями в каждой гамете | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| митоз | возникает только в том случае, если необходимо восстановить повреждение ДНК; обычно встречается между идентичными сестринскими хроматидами и не приводит к генетическим изменениям | ||||||||||
| число хромосом (плоидность) | мейоз | производит четыре генетически уникальные клетки, каждая с вдвое меньшим количеством хромосом, чем в родительской | |||||||||
| митоз | производит две генетически идентичные клетки, каждая с тем же числом хромосом, что и в родительской | ||||||||||
Мейоз начинается с диплоидной клетки, которая содержит две копии каждой хромосомы, называемые гомологами. Сначала клетка подвергается репликации ДНК, поэтому каждый гомолог теперь состоит из двух идентичных сестринских хроматид. Затем каждый набор гомологов соединяется друг с другом и обменивается генетической информацией посредством гомологичной рекомбинации, часто приводящей к физическим связям (кроссоверам) между гомологами. При первом делении мейоза гомологи разделяются на дочерние клетки с помощью веретенообразного аппарата. Затем клетки переходят ко второму делению без промежуточного цикла репликации ДНК. Сестринские хроматиды разделяются на дочерние клетки с образованием в общей сложности четырех гаплоидных клеток. Самки животных используют небольшую вариацию этого паттерна и производят одну большую яйцеклетку и два маленьких полярных тела. Из-за рекомбинации индивидуальная хроматида может состоять из новой комбинации материнской и отцовской генетической информации, в результате чего потомство генетически отличается от любого из родителей. Кроме того, индивидуальная гамета может включать в себя набор материнских, отцовских и рекомбинантных хроматид. Это генетическое разнообразие, возникающее в результате полового размножения, вносит свой вклад в разнообразие признаков, на которые может воздействовать естественный отбор.
Мейоз использует многие из тех же механизмов, что и митоз, тип деления клеток, используемый эукариотами для деления одной клетки на две идентичные дочерние клетки. У некоторых растений, грибов и протистов мейоз приводит к образованию спор : гаплоидных клеток, которые могут делиться вегетативно без оплодотворения. Некоторые эукариоты, такие как бделлоидные коловратки, не обладают способностью осуществлять мейоз и приобрели способность воспроизводиться путем партеногенеза.
Мейоз не встречается у архей или бактерий, которые обычно размножаются бесполым путем посредством бинарного деления. Однако «половой» процесс, известный как горизонтальный перенос генов, включает перенос ДНК от одной бактерии или археи к другой и рекомбинацию этих молекул ДНК различного родительского происхождения.
Мейоз был впервые обнаружен и описан в яйцах морских ежей в 1876 году немецким биологом Оскаром Хертвигом. Он был снова описан в 1883 году на уровне хромосом бельгийским зоологом Эдуардом Ван Бенеденом в яйцах аскарид аскарид. Однако значение мейоза для воспроизводства и наследования было описано только в 1890 году немецким биологом Августом Вейсманном, который отметил, что для сохранения количества хромосом необходимо два деления клеток, чтобы превратить одну диплоидную клетку в четыре гаплоидных. В 1911 году американский генетик Томас Хант Морган обнаружил кроссоверы в мейозе у плодовой мухи Drosophila melanogaster, что помогло установить, что генетические признаки передаются по хромосомам.
Термин «мейоз» происходит от греческого слова μείωσις, что означает «уменьшение». Он был введен в биологию Дж. Б. Фармером и Дж. Э. Муром в 1905 году с использованием своеобразного перевода «майоз»:
Мы предлагаем применять термины майоз или майотическая фаза для охвата всей серии ядерных изменений, включенных в два подразделения, которые Флемминг обозначил как гетеротип и гомотип.
Правописание было изменено на «мейоз» Кёрнике (1905) и Пантелем и Де Синети (1906), чтобы следовать обычным правилам транслитерации греческого языка.
Мейоз делится на мейоз I и мейоз II, которые далее подразделяются на кариокинез I и цитокинез I, а также кариокинез II и цитокинез II соответственно. Подготовительные шаги, которые приводят к мейозу, идентичны по структуре и названию интерфазе митотического клеточного цикла. Интерфаза разделена на три фазы:
За интерфазой следует мейоз I, затем мейоз II. Мейоз I разделяет реплицированные гомологичные хромосомы, каждая из которых по-прежнему состоит из двух сестринских хроматид, на две дочерние клетки, таким образом уменьшая число хромосом вдвое. Во время мейоза II сестринские хроматиды отделяются и образующиеся дочерние хромосомы разделяются на четыре дочерние клетки. У диплоидных организмов дочерние клетки, полученные в результате мейоза, являются гаплоидными и содержат только одну копию каждой хромосомы. У некоторых видов клетки входят в фазу покоя, известную как интеркинез между мейозом I и мейозом II.
Каждый мейоз I и II делится на профазную, метафазную, анафазную и телофазную стадии, сходные по назначению с их аналогичными субфазами в митотическом клеточном цикле. Следовательно, мейоз включает стадии мейоза I (профаза I, метафаза I, анафаза I, телофаза I) и мейоза II (профаза II, метафаза II, анафаза II, телофаза II).
 Схема мейотических фаз
Схема мейотических фаз Во время мейоза специфические гены транскрибируются в большей степени. В дополнение к сильной экспрессии мРНК, специфичной для мейотической стадии, существуют также повсеместные средства контроля трансляции (например, избирательное использование предварительно сформированной мРНК), регулирующие конечную мейотическую стадию специфической экспрессии белков генов во время мейоза. Таким образом, и транскрипционный, и трансляционный контроль определяют широкую реструктуризацию мейотических клеток, необходимую для осуществления мейоза.
Мейоз I разделяет гомологичные хромосомы, которые объединяются в тетрады (2n, 4c), производя две гаплоидные клетки (n хромосом, 23 у человека), каждая из которых содержит пары хроматид (1n, 2c). Поскольку плоидность снижается от диплоидной к гаплоидной, мейоз I называют редукционным делением. Мейоз II представляет собой эквациональное деление, аналогичное митозу, в котором сестринские хроматиды разделяются, создавая четыре гаплоидных дочерних клетки (1n, 1c).
 Профаза I мейоза у мышей. В Leptotene (L) начинают формироваться осевые элементы (окрашенные SYCP3). В зиготене (Z) поперечные элементы (SYCP1) и центральные элементы синаптонемного комплекса частично установлены (отображаются желтым цветом, поскольку они перекрываются с SYCP3). В пахитене (P) он полностью установлен, за исключением половых хромосом. В Diplotene (D) он разбирает раскрывающиеся хиазмы. КРЕСТ отмечает центромеры.
Профаза I мейоза у мышей. В Leptotene (L) начинают формироваться осевые элементы (окрашенные SYCP3). В зиготене (Z) поперечные элементы (SYCP1) и центральные элементы синаптонемного комплекса частично установлены (отображаются желтым цветом, поскольку они перекрываются с SYCP3). В пахитене (P) он полностью установлен, за исключением половых хромосом. В Diplotene (D) он разбирает раскрывающиеся хиазмы. КРЕСТ отмечает центромеры.  Схема синаптонемного комплекса на разных стадиях профазы I и хромосом, расположенных в виде линейного массива петель.
Схема синаптонемного комплекса на разных стадиях профазы I и хромосом, расположенных в виде линейного массива петель. Профаза I на сегодняшний день является самой продолжительной фазой мейоза (у мышей длится 13 из 14 дней). Во время профазы I гомологичные материнские и отцовские хромосомы объединяются в пары, синапсы и обмениваются генетической информацией (путем гомологичной рекомбинации ), образуя по крайней мере один кроссовер на хромосому. Эти пересечения становятся видимыми как хиазмы (множественное число; единственное число хиазмы ). Этот процесс способствует стабильному спариванию между гомологичными хромосомами и, следовательно, обеспечивает точное разделение хромосом при первом делении мейоза. Парные и реплицированные хромосомы называются бивалентами (две хромосомы) или тетрадами (четыре хроматиды ), причем по одной хромосоме от каждого родителя. Профаза I делится на ряд подстадий, названных в соответствии с внешним видом хромосом.
Первая стадия профазы I - это стадия лептотены, также известная как лептонема, от греческого слова, означающего «тонкие нити». На этой стадии профазы I отдельные хромосомы - каждая из которых состоит из двух реплицированных сестринских хроматид - становятся «индивидуализированными», образуя видимые нити внутри ядра. Каждая хромосома образует линейный массив петель, опосредованных когезином, а боковые элементы синаптонемного комплекса собираются, образуя «осевой элемент», из которого исходят петли. На этой стадии рекомбинация инициируется ферментом SPO11, который создает запрограммированные двухцепочечные разрывы (около 300 на мейоз у мышей). Этот процесс генерирует однонитевые нити ДНК, покрытые RAD51 и DMC1, которые вторгаются в гомологичные хромосомы, образуя межосевые мосты и приводя к спариванию / ко-выравниванию гомологов (на расстоянии ~ 400 нм у мышей).
За лептотеной следует стадия зиготены, также известная как зигонема, от греческих слов, означающих «парные нити», которые у некоторых организмов также называют стадией букета из-за того, как теломеры группируются на одном конце ядра. На этой стадии гомологичные хромосомы становятся намного более близкими (~ 100 нм) и стабильно спаренными (процесс, называемый синапсисом), опосредованный установкой поперечных и центральных элементов синаптонемного комплекса. Считается, что синапсис происходит по типу «застежки-молнии», начиная с узелка рекомбинации. Парные хромосомы называются двухвалентными или тетрадными хромосомами.
Пахитены этап ( / р æ к ɪ т я п / ПАК -i-подростков ), также известный как pachynema, от греческих слов, означающих «толстых нитей». это стадия, на которой все аутосомные хромосомы синапсируются. На этой стадии гомологичная рекомбинация, включая хромосомный кроссовер (кроссинговер), завершается за счет репарации двухцепочечных разрывов, образованных в лептотене. Большинство разрывов восстанавливается без образования кроссоверов, приводящих к конверсии генов. Однако подмножество разрывов (по крайней мере, по одному на хромосому) образуют кроссоверы между несестринскими (гомологичными) хромосомами, что приводит к обмену генетической информацией. Половые хромосомы, однако, не полностью идентичны и обмениваются информацией только в небольшой области гомологии, называемой псевдоавтосомной областью. Обмен информацией между гомологичными хроматидами приводит к рекомбинации информации; каждая хромосома имеет полный набор информации, которая у нее была раньше, и в результате этого процесса нет пробелов. Поскольку хромосомы не могут быть различимы в синаптонемном комплексе, фактический акт кроссинговера не воспринимается с помощью обычного светового микроскопа, а хиазмы не видны до следующей стадии.
Во время стадии диплотены, также известной как диплонема, от греческих слов, означающих «две нити», синаптонемный комплекс распадается, и гомологичные хромосомы немного отделяются друг от друга. Однако гомологичные хромосомы каждого бивалента остаются прочно связанными в хиазмах, областях, где произошел кроссинговер. Хиазмы остаются на хромосомах до тех пор, пока они не разрываются при переходе в анафазу I, чтобы позволить гомологичным хромосомам перемещаться к противоположным полюсам клетки.
В оогенезе плода человека все развивающиеся ооциты развиваются до этой стадии и задерживаются в профазе I до рождения. Это приостановленное состояние называется стадией диктиотена или диктиатом. Он длится до возобновления мейоза, чтобы подготовить ооцит к овуляции, которая происходит в период полового созревания или даже позже.
Хромосомы конденсируются дальше во время стадии диакинеза, от греческого слова, означающего «продвижение». Это первая точка мейоза, где фактически видны четыре части тетрад. Участки кроссинговера переплетаются друг с другом, эффективно перекрываясь, делая хиазмы четко видимыми. Помимо этого наблюдения, остальная часть стадии очень напоминает прометафазу митоза; в ядрышки исчезают, ядерная оболочка распадается в пузырьки, и мейоза шпинделя начинает формироваться.
В отличие от митотических клеток, ооциты человека и мыши не имеют центросом для образования мейотического веретена. У мышей примерно 80 центров организации микротрубочек (MTOC) образуют сферу в ооплазме и начинают зародышеобразование микротрубочек, которые достигают хромосом и прикрепляются к хромосомам на кинетохоре. Со временем MTOC сливаются, пока не образуются два полюса, образуя шпиндель бочкообразной формы. В человеческих ооцитах зарождение веретенообразных микротрубочек начинается на хромосомах, образуя звездочку, которая в конечном итоге расширяется и окружает хромосомы. Затем хромосомы скользят по микротрубочкам к экватору веретена, и в этот момент кинетохоры хромосом образуют концевые прикрепления к микротрубочкам.
Гомологичные пары движутся вместе вдоль метафазной пластинки: поскольку микротрубочки кинетохор от обоих полюсов веретена прикрепляются к своим соответствующим кинетохорам, парные гомологичные хромосомы выравниваются вдоль экваториальной плоскости, которая делит веретено пополам, из-за непрерывных уравновешивающих сил, действующих на биваленты микротрубочками, исходящими из две кинетохоры гомологичных хромосом. Эта насадка называется биполярной. Физической основой независимого набора хромосом является случайная ориентация каждого бивалента вдоль метафазной пластинки относительно ориентации других бивалентов вдоль той же экваториальной линии. Белковый комплекс когезин удерживает сестринские хроматиды вместе с момента их репликации до анафазы. В митозе сила вытягивания микротрубочек кинетохор в противоположных направлениях создает напряжение. Клетка ощущает это напряжение и не продвигается вперед с анафазой, пока все хромосомы не станут должным образом двунаправленными. В мейозе для установления напряжения обычно требуется по крайней мере один кроссовер на пару хромосом в дополнение к cohesin между сестринскими хроматидами (see Chromosome segregation ).
Микротрубочки кинетохор укорачиваются, притягивая гомологичные хромосомы (каждая из которых состоит из пары сестринских хроматид) к противоположным полюсам. Некинетохорные микротрубочки удлиняются, раздвигая центросомы дальше друг от друга. Клетка удлиняется, готовясь к делению по центру. В отличие от митоза, разрушается только когезин хромосомных плеч, в то время как когезин, окружающий центромеру, остается защищенным белком, названным Shugoshin (по-японски «дух-хранитель»), что препятствует разделению сестринских хроматид. Это позволяет сестринским хроматидам оставаться вместе, в то время как гомологи разделены.
Первое мейотическое деление эффективно заканчивается, когда хромосомы достигают полюсов. Каждая дочерняя клетка теперь имеет половину числа хромосом, но каждая хромосома состоит из пары хроматид. Микротрубочки, составляющие сеть веретена, исчезают, и каждый набор гаплоидов окружает новая ядерная мембрана. Хромосомы раскручиваются обратно в хроматин. Цитокинез, защемление клеточной мембраны в клетках животных или формирование клеточной стенки в клетках растений, завершается образованием двух дочерних клеток. Однако цитокинез не завершается полностью, что приводит к образованию «цитоплазматических мостов», которые позволяют разделять цитоплазму между дочерними клетками до конца мейоза II. Сестринские хроматиды остаются прикрепленными во время телофазы I.
Клетки могут войти в период покоя, известный как интеркинез или интерфаза II. На этом этапе репликации ДНК не происходит.
Мейоз II является вторым мейотическим делением и обычно включает эквациональную сегрегацию или разделение сестринских хроматид. Механически этот процесс похож на митоз, хотя его генетические результаты принципиально отличаются. Конечным результатом является производство четырех гаплоидных клеток (n хромосом, 23 у человека) из двух гаплоидных клеток (с n хромосомами, каждая из которых состоит из двух сестринских хроматид), образующихся в мейозе I. Четыре основных этапа мейоза II: профаза II., метафаза II, анафаза II и телофаза II.
В профазе II мы снова видим исчезновение ядрышек и ядерной оболочки, а также укорачивание и утолщение хроматид. Центросомы перемещаются в полярные регионы и организуют волокна веретена для второго деления мейоза.
В метафазе II центромеры содержат две кинетохоры, которые прикрепляются к волокнам веретена от центросом на противоположных полюсах. Новая экваториальная метафазная пластинка повернута на 90 градусов по сравнению с мейозом I, перпендикулярно предыдущей пластине.
За этим следует анафаза II, в которой оставшийся центромерный cohesin, больше не защищенный Shugoshin, расщепляется, позволяя сестринским хроматидам сегрегировать. Сестринские хроматиды по соглашению теперь называются сестринскими хромосомами, поскольку они движутся к противоположным полюсам.
Процесс заканчивается телофазой II, которая похожа на телофазу I, и отмечена деконденсацией и удлинением хромосом и разборкой веретена. Ядерные оболочки реформируются и отщепляются, или образование клеточной пластинки в конечном итоге дает в общей сложности четыре дочерних клетки, каждая с гаплоидным набором хромосом.
Теперь мейоз завершен и приводит к появлению четырех новых дочерних клеток.
Происхождение и функция мейоза в настоящее время не очень хорошо понимал, с научной точки зрения, и обеспечит фундаментальное понимание эволюции полового размножения в эукариот. В настоящее время среди биологов нет единого мнения по поводу того, как секс у эукариот возник в процессе эволюции, какую основную функцию выполняет половое размножение и почему оно сохраняется, учитывая основную двукратную стоимость секса. Ясно, что он появился более 1,2 миллиарда лет назад, и что почти все виды, являющиеся потомками первоначальных видов, воспроизводящих половым путем, по-прежнему являются половыми размножителями, включая растения, грибы и животных.
Мейоз - ключевое событие полового цикла эукариот. Это стадия жизненного цикла, когда клетка дает начало гаплоидным клеткам ( гаметам ), каждая из которых имеет вдвое меньше хромосом, чем родительская клетка. Две такие гаплоидные гаметы, обычно возникающие из разных индивидуальных организмов, сливаются в процессе оплодотворения, завершая, таким образом, половой цикл.
Мейоз распространен среди эукариот. Это происходит в одноклеточных организмах, таких как дрожжи, а также в многоклеточных организмах, таких как люди. Эукариоты произошли от прокариот более 2,2 миллиарда лет назад, и самые ранние эукариоты, вероятно, были одноклеточными организмами. Чтобы понять пол у эукариот, необходимо понять (1) как возник мейоз у одноклеточных эукариот и (2) функция мейоза.
Новые комбинации ДНК, созданные во время мейоза, являются значительным источником генетических вариаций наряду с мутациями, что приводит к новым комбинациям аллелей, которые могут быть полезными. Мейоз порождает генетическое разнообразие гамет двумя способами: (1) Закон независимого ассортимента. Независимая ориентация гомологичных пар хромосом вдоль метафазной пластинки во время метафазы I и ориентация сестринских хроматид в метафазе II, это последующее разделение гомологов и сестринских хроматид во время анафазы I и II, это позволяет случайное и независимое распределение хромосом по каждой из них. дочерняя клетка (и, в конечном итоге, до гамет); и (2) переход. Физический обмен гомологичных участков хромосом путем гомологичной рекомбинации во время профазы I приводит к новым комбинациям генетической информации в хромосомах.
Самки млекопитающих и птиц рождаются со всеми ооцитами, необходимыми для будущих овуляций, и эти ооциты задерживаются на профазе I стадии мейоза. У людей, например, ооциты образуются между тремя и четырьмя месяцами беременности внутри плода и, следовательно, присутствуют при рождении. Во время этой арестованной стадии профазы I ( диктиата ), которая может длиться десятилетиями, в ооцитах присутствуют четыре копии генома. Арест ooctyes на этапе копирования четыре генома был предложен обеспечить информационную избыточность, необходимую для ремонта повреждений в ДНК в зародышевом. Используемый процесс репарации, по-видимому, включает гомологичную рекомбинационную репарацию. Ооциты, задержанные в профазе I, обладают высокой способностью к эффективной репарации повреждений ДНК, особенно экзогенно индуцированных двухцепочечных разрывов. Способность к репарации ДНК, по-видимому, является ключевым механизмом контроля качества в женской зародышевой линии и критическим детерминантом фертильности.
 Дипломированный жизненный цикл
Дипломированный жизненный цикл  Гаплонтический жизненный цикл. Основная статья: Биологический жизненный цикл Смотрите также: Смена поколений
Гаплонтический жизненный цикл. Основная статья: Биологический жизненный цикл Смотрите также: Смена поколений Мейоз возникает в жизненных циклах эукариот, включающих половое размножение, состоящее из постоянного циклического процесса мейоза и оплодотворения. Это происходит вместе с нормальным делением митотических клеток. В многоклеточных организмах существует промежуточный этап между диплоидным и гаплоидным переходом, на котором организм растет. На определенных этапах жизненного цикла половые клетки производят гаметы. Соматические клетки составляют тело организма и не участвуют в производстве гамет.
Чередование событий мейоза и оплодотворения вызывает серию переходов назад и вперед между чередующимися гаплоидными и диплоидными состояниями. Фаза жизненного цикла организма может происходить либо во время диплоидного состояния ( диплонтический жизненный цикл), либо во время гаплоидного состояния ( гаплонтический жизненный цикл), либо во время обоих ( гаплодиплонический жизненный цикл, в котором есть две отдельные фазы организма, одна во время гаплоидного цикла). состояние, а другой - в диплоидном состоянии). В этом смысле существует три типа жизненных циклов, в которых используется половое размножение, которые различаются расположением фазы (фаз) организма.
В диплоидном жизненном цикле (с предгаметическим мейозом), частью которого являются люди, организм является диплоидным, выросшим из диплоидной клетки, называемой зиготой. Диплоидные стволовые клетки зародышевой линии организма подвергаются мейозу, чтобы создать гаплоидные гаметы ( сперматозоиды для мужчин и яйцеклетки для женщин), которые оплодотворяются, образуя зиготу. Диплоидная зигота подвергается многократному клеточному делению путем митоза и врастает в организм.
В гаплонтическом жизненном цикле (с постзиготическим мейозом) организм гаплоидный, порожденный пролиферацией и дифференцировкой единственной гаплоидной клетки, называемой гаметой. Два организма противоположного пола вносят свои гаплоидные гаметы в диплоидную зиготу. Зигота немедленно подвергается мейозу, образуя четыре гаплоидных клетки. Эти клетки подвергаются митозу, чтобы создать организм. Многие грибы и многие простейшие используют гаплонтический жизненный цикл.
Наконец, в гаплодипонтическом жизненном цикле (со споровым или промежуточным мейозом) живой организм чередует гаплоидные и диплоидные состояния. Следовательно, этот цикл также известен как смена поколений. Клетки зародышевой линии диплоидного организма подвергаются мейозу с образованием спор. Споры размножаются митозом, превращаясь в гаплоидный организм. Затем гамета гаплоидного организма объединяется с гаметой другого гаплоидного организма, создавая зиготу. Зигота подвергается многократному митозу и дифференцировке, чтобы снова стать диплоидным организмом. Гаплодипонтический жизненный цикл можно рассматривать как сплав диплонтического и гаплонтического жизненных циклов.
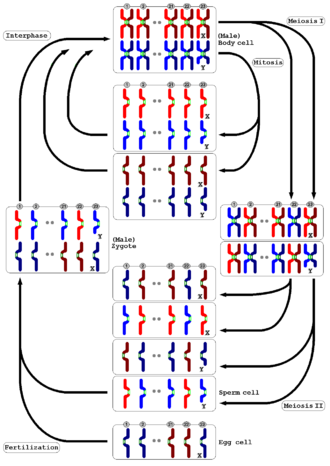 Обзор распределения хроматид и хромосом в митотическом и мейотическом цикле мужской клетки человека
Обзор распределения хроматид и хромосом в митотическом и мейотическом цикле мужской клетки человека Мейоз встречается у всех животных и растений. Конечный результат, производство гамет с половиной количества хромосом по сравнению с родительской клеткой, такой же, но подробный процесс отличается. У животных мейоз непосредственно производит гаметы. У наземных растений и некоторых водорослей происходит чередование поколений, так что мейоз в поколении диплоидных спорофитов дает гаплоидные споры. Эти споры размножаются путем митоза, развиваясь в поколение гаплоидных гаметофитов, которые затем непосредственно дают начало гаметам (то есть без дальнейшего мейоза). И у животных, и у растений заключительный этап - слияние гамет, восстанавливающее исходное количество хромосом.
У женщин мейоз происходит в клетках, известных как ооциты (единственное число: ооциты ). Каждый первичный ооцит делится дважды в мейозе, в каждом случае неравномерно. Первое деление дает дочернюю клетку и полярное тело гораздо меньшего размера, которое может или не может подвергаться второму делению. В мейозе II деление дочерней клетки дает второе полярное тело и единственную гаплоидную клетку, которая увеличивается в размерах и становится яйцеклеткой. Следовательно, у самок каждый первичный ооцит, претерпевающий мейоз, дает одну зрелую яйцеклетку и одно или два полярных тельца.
Обратите внимание, что у самок во время мейоза бывают паузы. Созревающие ооциты задерживаются в профазе I мейоза I и бездействуют в защитной оболочке соматических клеток, называемой фолликулом. В начале каждого менструального цикла, FSH секреция из передней доли гипофиза стимулирует несколько фолликулов созревает в процессе, известном как фолликулогенез. Во время этого процесса созревающие ооциты возобновляют мейоз и продолжаются до метафазы II мейоза II, где они снова задерживаются непосредственно перед овуляцией. Если эти ооциты оплодотворены спермой, они возобновят и завершат мейоз. Во время фолликулогенеза у человека обычно один фолликул становится доминирующим, в то время как другие подвергаются атрезии. Процесс мейоза у самок происходит во время оогенеза и отличается от типичного мейоза тем, что он имеет длительный период мейотического ареста, известный как стадия диктиата, и лишен помощи центросом.
У мужчин, мейоз происходит во время сперматогенеза в семенных канальцах этих яичек. Мейоз во время сперматогенеза специфичен для типа клеток, называемых сперматоцитами, которые позже созреют и станут сперматозоидами. Мейоз первичных половых клеток происходит в период полового созревания, намного позже, чем у женщин. Ткани мужских семенников подавляют мейоз за счет разложения ретиноевой кислоты, которая, как предполагается, является стимулятором мейоза. Это преодолевается в период полового созревания, когда клетки семенных канальцев, называемые клетками Сертоли, начинают вырабатывать собственную ретиноевую кислоту. Чувствительность к ретиноевой кислоте также регулируется белками, называемыми нано и DAZL. Исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота необходима постнатально для стимуляции дифференцировки сперматогониев, которая через несколько дней проявляется в сперматоцитах, подвергающихся мейозу, однако ретиноевая кислота не требуется во время инициации мейоза.
У самок млекопитающих мейоз начинается сразу после миграции первичных половых клеток в яичник эмбриона. Некоторые исследования предполагают, что ретиноевая кислота, полученная из примитивной почки (мезонефрос), стимулирует мейоз в эмбриональных яичниковых оогониях и что ткани эмбриональных мужских семенников подавляют мейоз, разлагая ретиноевую кислоту. Однако исследования генетической потери функции ферментов, генерирующих ретиноевую кислоту, показали, что ретиноевая кислота не требуется ни для инициации женского мейоза, который происходит во время эмбриогенеза, ни для мужского мейоза, который инициируется постнатально.
В то время как большинство эукариот имеют мейоз с двумя делениями (хотя иногда и ахиазматический ), очень редкая форма, мейоз с одним делением, встречается у некоторых жгутиконосцев ( парабазалид и оксимонад ) из кишечника питающегося деревом таракана Cryptocercus.
Рекомбинация между 23 парами хромосом человека отвечает за перераспределение не только самих хромосом, но и частей каждой из них. Также, согласно оценкам, рекомбинация у самок в 1,6 раза больше, чем у самцов. Кроме того, в среднем женская рекомбинация выше на центромерах, а мужская рекомбинация выше на теломерах. В среднем 1 миллион пар оснований (1 МБ) соответствует 1 сМоргану (см = 1% частота рекомбинации). Частота кроссоверов остается неопределенной. У дрожжей, мышей и человека было подсчитано, что в пермейотической клетке образуется ≥200 двунитевых разрывов (DSB). Однако только подмножество DSB (~ 5–30% в зависимости от организма) продолжает давать кроссоверы, которые приводят только к 1-2 кроссоверам на хромосому человека.
Нормальное разделение хромосом в мейозе I или сестринских хроматид в мейозе II называется дизъюнкцией. Когда сегрегация не является нормальной, это называется нерасхождением. Это приводит к образованию гамет, которые имеют либо слишком много, либо слишком мало определенной хромосомы, и является обычным механизмом для трисомии или моносомии. Нерасхождение может происходить в мейозе I или мейозе II, фазах клеточного воспроизводства или во время митоза.
Большинство моносомных и трисомных человеческих эмбрионов нежизнеспособны, но некоторые анеуплоидии могут переноситься, например трисомия по наименьшей хромосоме, хромосоме 21. Фенотипы этих анеуплоидий варьируются от тяжелых нарушений развития до бессимптомных. Медицинские условия включают, но не ограничиваются:
Вероятность нерасхождения в человеческих ооцитах увеличивается с увеличением возраста матери, предположительно из-за потери когезина с течением времени.
Для понимания мейоза полезно сравнение с митозом. В таблице ниже показаны различия между мейозом и митозом.
| Мейоз | Митоз | |
|---|---|---|
| Конечный результат | Обычно четыре клетки, каждая с половиной хромосом в каждой из них. | Две клетки, имеющие такое же количество хромосом, что и родительская |
| Функция | Производство гамет (половых клеток) у эукариот, размножающихся половым путем, с диплонтовым жизненным циклом | Клеточное размножение, рост, ремонт, бесполое размножение |
| Где это происходит? | Почти все эукариоты (животные, растения, грибы и простейшие ); В гонадах, перед гаметами (в диплонтических жизненных циклах); После зигот (в гаплонтике); Перед спорами (при гаплодиплонтике) | Все пролиферирующие клетки у всех эукариот |
| Шаги | Профаза I, Метафаза I, Анафаза I, Телофаза I, Профаза II, Метафаза II, Анафаза II, Телофаза II | Профаза, Прометафаза, Метафаза, Анафаза, Телофаза |
| Генетически такой же, как родитель? | Нет | да |
| Переход происходит? | Да, обычно встречается между каждой парой гомологичных хромосом | Очень редко |
| Спаривание гомологичных хромосом? | да | Нет |
| Цитокинез | Встречается в телофазе I и телофазе II. | Встречается в телофазе |
| Центромеры расщепляются | Не встречается в Анафазе I, но встречается в Анафазе II. | Происходит в анафазе |
Как клетка переходит к мейотическому делению при мейотическом делении клеток, хорошо не известно. Фактор, способствующий созреванию (MPF), по- видимому, играет роль в мейозе ооцитов лягушки. У гриба S. pombe. Связывающий белок MeiRNA участвует в делении мейотических клеток.
Было высказано предположение, что продукт гена CEP1 дрожжей, который связывает центромерную область CDE1, может играть роль в спаривании хромосом во время мейоза-I.
Мейотическая рекомбинация опосредуется двухцепочечным разрывом, который катализируется белком Spo11. Также Mre11, Sae2 и Exo1 играют роль в разрыве и рекомбинации. После разрыва происходит рекомбинация, которая обычно гомологична. Рекомбинация может происходить либо по пути двойного соединения Холлидея (dHJ), либо по пути зависимого от синтеза отжига цепи (SDSA). (Второй относится к непересекающемуся продукту).
Похоже, есть контрольные точки и для деления мейотических клеток. Считается, что в S. pombe белки Rad, S. pombe Mek1 (с киназным доменом FHA), Cdc25, Cdc2 и неизвестный фактор образуют контрольную точку.
В оогенезе позвоночных, поддерживаемый цитостатическим фактором (CSF), играет роль в переключении на мейоз-II.