
Войти
| Neurospora crassa | |
|---|---|
 | |
| Научная классификация | |
| Царство: | Грибы |
| Тип: | Ascomycota |
| Подтип: | Pezizomycotina |
| Класс: | Sordariomycetes |
| Порядок: | Sordariales |
| Семейство: | Sordariaceae |
| Род: | Neurospora |
| Виды: | N. crassa |
| Биномиальное имя | |
| Neurospora crassa . Shear B.O. Dodge | |
Neurospora crassa - это разновидность плесени для красного хлеба типа Ascomycota. Название рода, означающее по-гречески «нервная спора», относится к характерным бороздкам на спорах. Первое опубликованное сообщение об этом грибке было связано с заражением французских пекарен в 1843 году.
N. crassa используется в качестве модельного организма, потому что он легко выращивается и имеет гаплоидный жизненный цикл, что упрощает генетический анализ, поскольку рецессивные признаки будут обнаруживаться в потомство. Анализ генетической рекомбинации облегчается упорядоченным расположением продуктов мейоза в аскоспорах Neurospora . Его весь геном из семи хромосом был секвенирован.
Neurospora использовалась Эдвардом Татумом и Джорджем Уэллсом Бидлом в своих экспериментах, для которых они получил Нобелевскую премию по физиологии и медицине в 1958 году. Бидл и Татум подвергли N. crassa рентгеновскому излучению, вызвав мутации. Затем они наблюдали сбои в метаболических путях, вызванные ошибками в конкретных ферментах. Это привело их к предложению гипотезы «один ген, один фермент», согласно которой конкретные гены кодируют определенные белки. Их гипотеза была позже развита для ферментных путей Норманом Горовицем, также работавшим над Neurospora. Как вспоминал Норман Горовиц в 2004 году: «Эти эксперименты положили начало науке о том, что Бидл и Татум назвали« биохимической генетикой ». На самом деле они оказались первым оружием в том, что стало молекулярной генетикой и всеми последующими разработками».
В выпуске Nature от 24 апреля 2003 г. геном N. crassa был описан как полностью секвенированный. Геном имеет длину около 43 мегабаз и включает примерно 10 000 генов. Осуществляется проект по производству штаммов, содержащих нокаутные мутанты каждого гена N. crassa.
В естественной среде N. crassa обитает в основном в тропических и субтропических регионах. Его можно найти растущим на мертвых растениях после пожаров.
Neurospora активно используется в исследованиях по всему миру. Это важно для выяснения молекулярных событий, связанных с циркадными ритмами, эпигенетикой и подавлением генов, полярностью клеток, клетками. слияние, развитие, а также многие аспекты клеточной биологии и биохимии.
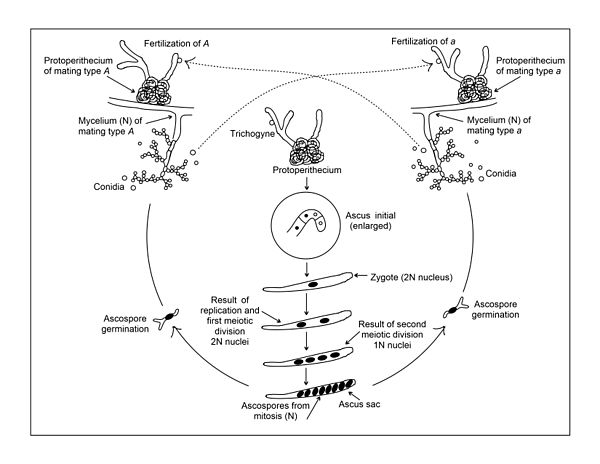 Жизненный цикл Neurospora crassa. Гаплоидный мицелий воспроизводится бесполым путем посредством двух процессов: (1) простой пролиферации существующего мицелия и (2) образования конидий (макро- и микро-), которые могут диспергироваться, а затем прорастать с образованием нового мицелия. В половом цикле спаривание может происходить только между отдельными линиями разного типа спаривания, A и a. Оплодотворение происходит путем прохождения ядер конидий или мицелия одного типа спаривания в протоперитеции противоположного типа через трихогин. Слияние ядер противоположных типов спаривания происходит внутри протоперитеция с образованием ядра зиготы (2N).
Жизненный цикл Neurospora crassa. Гаплоидный мицелий воспроизводится бесполым путем посредством двух процессов: (1) простой пролиферации существующего мицелия и (2) образования конидий (макро- и микро-), которые могут диспергироваться, а затем прорастать с образованием нового мицелия. В половом цикле спаривание может происходить только между отдельными линиями разного типа спаривания, A и a. Оплодотворение происходит путем прохождения ядер конидий или мицелия одного типа спаривания в протоперитеции противоположного типа через трихогин. Слияние ядер противоположных типов спаривания происходит внутри протоперитеция с образованием ядра зиготы (2N). Половые плодовые тела (перитеции) могут образовываться только тогда, когда встречаются два мицелия разного типа спаривания (см. Рисунок). Как и другие аскомицеты, N. crassa имеет два типа спаривания, которые в данном случае обозначаются буквами A и a. Явных морфологических различий между штаммами типа А и а не наблюдается. Оба могут образовывать обильные протоперитеции, женскую репродуктивную структуру (см. Рисунок). Протоперитеции легче всего образуются в лаборатории, когда рост происходит на твердой (агаровой) синтетической среде с относительно низким источником азота. По-видимому, азотное голодание необходимо для экспрессии генов, участвующих в половом развитии. Протоперитеций состоит из аскогониума, спиральной многоклеточной гифы, заключенной в узелковое скопление гиф. Разветвленная система тонких гиф, называемая трихогин, простирается от кончика аскогониума, выступая за покровные гифы в воздух. Половой цикл начинается (т.е. происходит оплодотворение), когда клетка (обычно конидий) противоположного типа спаривания контактирует с частью трихогина (см. Рисунок). За таким контактом может последовать слияние клеток, ведущее к одному или нескольким ядрам из оплодотворяющей клетки, мигрирующим вниз по трихогину в аскогониум. Поскольку оба штамма A и a имеют одинаковую половую структуру, ни один из них не может считаться исключительно мужским или женским. Однако в качестве реципиента протоперитеций штаммов A и a можно рассматривать как женскую структуру, а оплодотворяющий конидий можно рассматривать как участника мужского пола.
Последующие этапы после слияния А и гаплоидных клеток были описаны Финчемом и Дей, Вагнером и Митчеллом. После слияния клеток дальнейшее слияние их ядер задерживается. Вместо этого ядро оплодотворяющей клетки и ядро аскогониума связываются и начинают синхронно делиться. Продукты этих ядерных делений (все еще в парах разного типа спаривания, то есть A / a) мигрируют в многочисленные аскогенные гифы, которые затем начинают расти из аскогониума. Каждая из этих аскогенных гиф изгибается, образуя крючок (или крючок) на своем конце, а А и пара гаплоидных ядер внутри крючка разделяются синхронно. Затем формируются перегородки, чтобы разделить жердь на три ячейки. Центральная ячейка изгиба крючка содержит одно ядро А и одно ядро А (см. Рисунок). Эта биядерная клетка инициирует образование асков и называется «первичной аскусной» клеткой. Затем две одноядерные клетки по обе стороны от первой аскообразующей клетки сливаются друг с другом, образуя двухъядерную клетку, которая может расти, образуя еще одну клетку, которая затем может образовывать свою собственную аскус-начальная клетка. Затем этот процесс можно повторять несколько раз.
После образования клетки-аска, А и ядро сливаются друг с другом, образуя диплоидное ядро (см. Рисунок). Это ядро - единственное диплоидное ядро во всем жизненном цикле N. crassa. Диплоидное ядро состоит из 14 хромосом, образованных из двух слитых гаплоидных ядер, каждое из которых имеет по 7 хромосом. За образованием диплоидного ядра сразу следует мейоз. Два последовательных деления мейоза приводят к четырем гаплоидным ядрам, двум типу спаривания A и двум типу спаривания. Одно дальнейшее митотическое деление приводит к четырем А и четырем ядрам в каждой аске. Мейоз является неотъемлемой частью жизненного цикла всех организмов, размножающихся половым путем, и в своих основных чертах мейоз у N. crassa кажется типичным для мейоза в целом.
По мере того, как происходят вышеуказанные события, мицелиальная оболочка, которая окружала аскогоний, развивается как стенка перитеция, пропитывается меланином и почернеет. Зрелый перитеций имеет колбообразное строение.
Зрелый перитеций может содержать до 300 асков, каждый из которых происходит из идентичных диплоидных ядер слияния. Обычно в природе, когда перитеции созревают, аскоспоры довольно сильно выбрасываются в воздух. Эти аскоспоры термостойки и в лаборатории требуют нагревания при 60 ° C в течение 30 минут, чтобы вызвать прорастание. У нормальных штаммов весь половой цикл занимает от 10 до 15 дней. В зрелом аске, содержащем восемь аскоспор, пары соседних спор идентичны по генетической конституции, так как последнее деление является митотическим, и поскольку аскоспоры содержатся в мешочке асков, который удерживает их в определенном порядке, определяемом направлением ядерных сегрегаций во время мейоз. Поскольку четыре первичных продукта также расположены последовательно, паттерн сегрегации первого деления генетических маркеров можно отличить от паттерна сегрегации второго деления.
Из-за перечисленных выше особенностей N. crassa оказалась очень полезной для изучения генетических событий, происходящих в отдельных мейозах. Зрелые аски от перитеция можно отделить на предметном стекле микроскопа и экспериментально обработать спорами. Эти исследования обычно включали отдельную культуру отдельных аскоспор, полученных в результате одного мейотического события, и определение генотипа каждой споры. Исследования этого типа, проведенные в нескольких различных лабораториях, установили феномен «конверсии генов» (например, см. Ссылки).
В качестве примера феномена конверсии гена рассмотрим генетические скрещивания двух мутантных штаммов N. crassa, дефектных по гену pan-2. Этот ген необходим для синтеза пантотеновой кислоты (витамина B5), и мутанты, дефектные по этому гену, могут быть экспериментально идентифицированы по их потребности в пантотеновой кислоте в их питательной среде. Две мутации pan-2, B5 и B3, расположены в разных сайтах в гене pan-2, так что скрещивание B5 ´ B3 дает рекомбинанты дикого типа с низкой частотой. Анализ 939 асков, в котором можно было определить генотипы всех продуктов мейоза (аскоспоры), обнаружил 11 асков с исключительным паттерном сегрегации. Они включали шесть асков, в которых был один мейотический продукт дикого типа, но не было ожидаемого реципрокного продукта двойного мутанта (B5B3). Кроме того, в трех асках соотношение продуктов мейоза было 1B5: 3B3, а не ожидаемое соотношение 2: 2. Это исследование, а также многочисленные дополнительные исследования на N. crassa и других грибах (обзор Уайтхауса) привели к обширной характеристике конверсии генов. Из этой работы стало ясно, что события конверсии генов возникают, когда событие молекулярной рекомбинации происходит рядом с исследуемыми генетическими маркерами (например, мутации pan-2 в приведенном выше примере). Таким образом, исследования конверсии генов позволили понять детали молекулярного механизма рекомбинации. За десятилетия, прошедшие после первых наблюдений Мэри Митчелл в 1955 году, была предложена последовательность молекулярных моделей рекомбинации, основанная как на новых генетических данных, полученных в результате исследований конверсии генов, так и на исследованиях реакционных способностей ДНК. Текущее понимание молекулярного механизма рекомбинации обсуждается в статьях Википедии Конверсия генов и Генетическая рекомбинация. Понимание рекомбинации имеет отношение к нескольким фундаментальным биологическим проблемам, таким как роль рекомбинации и рекомбинационная репарация при раке (см. BRCA1 ) и адаптивная функция мейоза (см. Мейоз ).
То, что спаривание у N. crassa может происходить только между штаммами разного типа спаривания, предполагает, что естественный отбор способствует некоторой степени ауткроссинга. У гаплоидных многоклеточных грибов, таких как N. crassa, мейоз, протекающий на короткой диплоидной стадии, является одним из наиболее сложных процессов. Гаплоидная многоклеточная вегетативная стадия, хотя физически намного больше, чем диплоидная стадия, для нее характерна простая модульная конструкция с небольшой дифференциацией. У N. crassa рецессивные мутации, влияющие на диплоидную стадию жизненного цикла, довольно часто встречаются в естественных популяциях. Эти мутации, когда они гомозиготны на диплоидной стадии, часто вызывают дефекты созревания спор или образование бесплодных плодовых тел с небольшим количеством аскоспор (половых спор). Большинство этих гомозиготных мутаций вызывают аномальный мейоз (например, нарушение спаривания хромосом или нарушение пахитены или диплотены). Число генов, влияющих на диплоидную стадию, оценивалось как минимум 435 (около 4% от общего числа 9730 генов). Таким образом, ауткроссинг, вызванный необходимостью объединения противоположных типов спаривания, вероятно, обеспечивает преимущество маскировки рецессивных мутаций, которые в противном случае были бы вредны для образования половых спор (см. Комплементация (генетика) ).
Neurospora crassa - это не только модельный организм для изучения фенотипических типов в нокаутированных вариантах, но и особенно полезный организм, широко используемый в вычислительной биологии и циркадные часы. Его естественный репродуктивный цикл составляет 22 часа, и на него влияют внешние факторы, такие как свет и температура. Нокаут-варианты N. crassa дикого типа широко изучаются для определения влияния конкретных генов (см. Частота (ген) ).
| На Викискладе есть материалы, связанные с Neurospora crassa. |