
Войти
 Типичная структура зрелой эукариотической мРНК
Типичная структура зрелой эукариотической мРНК Полиаденилирование добавление поли (A) хвоста к информационной РНК. Поли (A) хвост состоит из множества аденозинмонофосфатов ; другими словами, это участок РНК, который имеет только аденин оснований. У эукариот полиаденилирование является частью процесса, который производит зрелую информационную РНК (мРНК) для трансляции. У многих бактерий поли (A) хвост способствует деградации мРНК. Следовательно, он составляет часть более крупного процесса экспрессии гена .
. Процесс полиаденилирования начинается, когда транскрипция гена заканчивается. 3'-крайний сегмент вновь образованной пре-мРНК сначала отщепляется набором белков ; эти белки затем синтезируют поли (А) хвост на 3'-конце РНК. В некоторых генах эти белки добавляют поли (А) хвост в одном из нескольких возможных сайтов. Следовательно, полиаденилирование может производить более одного транскрипта из одного гена (альтернативное полиаденилирование ), аналогично альтернативному сплайсингу.
Поли (A) -хвост важен для ядерного экспорта, трансляции и стабильность мРНК. Хвост со временем укорачивается, и, когда он достаточно короткий, мРНК подвергается ферментативной деградации. Однако в некоторых типах клеток мРНК с короткими поли (A) хвостами сохраняются для последующей активации путем повторного полиаденилирования в цитозоле. Напротив, когда полиаденилирование происходит у бактерий, оно способствует деградации РНК. Это также иногда случается с эукариотическими некодирующими РНК.
Молекулы мРНК как у прокариот, так и у эукариот имеют полиаденилированные 3'-концы, при этом прокариотические поли (A) хвосты обычно короче и меньше полиаденилированных молекул мРНК.
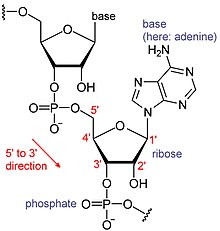 Химическая структура РНК. Последовательность оснований различается между молекулами РНК.
Химическая структура РНК. Последовательность оснований различается между молекулами РНК. РНК представляют собой тип больших биологических молекул, отдельные строительные блоки которых называются нуклеотидами. Название поли (A) хвост (для хвоста полиадениловой кислоты) отражает способ сокращения нуклеотидов РНК, с буквой для основания, которое содержит нуклеотид (A для аденин, C для цитозин, G для гуанина и U для урацила ). РНК продуцируются (транскрибируются ) из матрицы ДНК. По соглашению, последовательности РНК записываются в направлении от 5 'до 3'. 5'-конец - это часть молекулы РНК, которая транскрибируется первой, а 3'-конец транскрибируется последней. На 3'-конце также находится поли (A) хвост на полиаденилированных РНК.
Информационная РНК (мРНК) - это РНК, которая имеет кодирующую область, которая действует как матрица для синтеза белка (трансляция ). Остальная часть мРНК, нетранслируемые области, настраивает, насколько активна мРНК. Есть также много не транслируемых РНК, называемых некодирующими РНК. Подобно нетранслируемым областям, многие из этих некодирующих РНК выполняют регуляторные роли.
При ядерном полиаденилировании к поли (А) хвосту добавляется РНК в конце транскрипции. На мРНК поли (A) хвост защищает молекулу мРНК от ферментативной деградации в цитоплазме и помогает в терминации транскрипции, экспорте мРНК из ядра и трансляции. Почти все мРНК эукариот полиаденилированы, за исключением мРНК гистона животных, зависимых от репликации. Это единственные мРНК у эукариот, у которых отсутствует поли (A) -хвост, который вместо этого заканчивается структурой стебель-петля, за которой следует богатая пуринами последовательность, называемая гистоновым нижележащим элементом, который указывает, где разрезана РНК. так что образуется 3'-конец мРНК гистона.
Многие эукариотические некодирующие РНК всегда полиаденилируются в конце транскрипции. Существуют небольшие РНК, в которых поли (А) хвост виден только в промежуточных формах, а не в зрелой РНК, поскольку концы удаляются во время процессинга, наиболее заметными из которых являются микроРНК. Но для многих длинных некодирующих РНК - кажущаяся большая группа регуляторных РНК, которая, например, включает РНК Xist, которая опосредует Х-хромосому инактивация - поли (A) хвост является частью зрелой РНК.
| Участвующие белки:. CPSF : фактор специфичности расщепления / полиаденилирования. CstF : фактор стимуляции расщепления. PAP : полиаденилатполимераза. PABII : полиаденилатсвязывающий белок 2. CFI : фактор расщепления I. CFII : фактор расщепления II |
Комплекс процессивного полиаденилирования в ядре эукариот воздействует на продукты РНК-полимеразы II, например мРНК предшественника. Здесь мультибелковый комплекс (см. Компоненты справа) расщепляет 3'-большую часть вновь продуцируемой РНК и полиаденилирует конец, полученный в результате этого расщепления. Расщепление катализируется ферментом CPSF и происходит на 10–30 нуклеотидов ниже его сайта связывания. Этот сайт часто имеет сигнальную последовательность полиаденилирования AAUAAA на РНК, но существуют ее варианты, которые более слабо связываются с CPSF. Два других белка добавляют специфичность связыванию с РНК: CstF и CFI. CstF связывается с GU-богатой областью ниже по ходу от сайта CPSF. CFI распознает третий сайт на РНК (набор последовательностей UGUAA у млекопитающих) и может рекрутировать CPSF, даже если последовательность AAUAAA отсутствует. Сигнал полиаденилирования - мотив последовательности, распознаваемый комплексом расщепления РНК, - варьируется между группами эукариот. Большинство сайтов полиаденилирования человека содержат последовательность AAUAAA, но эта последовательность менее распространена у растений и грибов.
РНК обычно расщепляется до терминации транскрипции, поскольку CstF также связывается с РНК-полимеразой II. Благодаря плохо изученному механизму (по состоянию на 2002 г.) он дает сигнал РНК-полимеразе II, чтобы ускользнуть из транскрипта. В расщеплении также участвует белок CFII, но неизвестно, каким образом. Сайт расщепления, связанный с сигналом полиаденилирования, может варьироваться примерно до 50 нуклеотидов.
Когда РНК расщепляется, начинается полиаденилирование, катализируемое полиаденилатполимеразой. Полиаденилатполимераза формирует поли (A) хвост путем добавления аденозинмонофосфатных единиц из аденозинтрифосфата к РНК, отщепляя пирофосфат. Другой белок, PAB2, связывается с новым коротким поли (A) хвостом и увеличивает сродство полиаденилатполимеразы к РНК. Когда длина поли (A) хвоста составляет приблизительно 250 нуклеотидов, фермент больше не может связываться с CPSF, и полиаденилирование прекращается, определяя таким образом длину поли (A) хвоста. CPSF находится в контакте с РНК-полимеразой II, что позволяет ей сигнализировать полимеразе о прекращении транскрипции. Когда РНК-полимераза II достигает «терминирующей последовательности» ('TTTATT³ на матрице ДНК и ⁵'AAUAAA³ на первичном транскрипте), сигнализируется окончание транскрипции. Аппарат полиаденилирования также физически связан со сплайсосомой , комплексом, который удаляет интроны из РНК.
Поли (A) хвост действует как сайт связывания для поли (A) -связывающего белка. Поли (A) -связывающий белок способствует экспорту из ядра и трансляции, а также ингибирует деградацию. Этот белок связывается с поли (А) хвостом до экспорта мРНК из ядра, а в дрожжах также рекрутирует поли (А) нуклеазу, фермент, который укорачивает поли (А) хвост и позволяет экспортировать мРНК. Поли (A) -связывающий белок экспортируется в цитоплазму вместе с РНК. мРНК, которые не экспортируются, разрушаются экзосомой. Поли (A) -связывающий белок также может связываться с несколькими белками, которые влияют на трансляцию, и, таким образом, рекрутировать их, одним из них является фактор инициации -4G, который, в свою очередь, привлекает рибосомные 40S субъединица. Однако поли (А) хвост не требуется для трансляции всех мРНК. Кроме того, поли (A) хвостовая часть (олигоаденилирование) может определять судьбу молекул РНК, которые обычно не являются поли (A) -концевыми (такими как (небольшие) некодирующие (sn) РНК и т. Д.), И тем самым индуцировать их РНК.
В эукариотических соматических клетках поли (A) хвосты большинства мРНК в цитоплазме постепенно укорачиваются, а мРНК с более коротким поли (A) хвосты переводятся меньше и быстрее деградируют. Однако может пройти много часов, прежде чем мРНК распадется. Этот процесс деаденилирования и деградации может быть ускорен микроРНК, комплементарными 3'-нетранслируемой области мРНК. В незрелых яйцеклетках мРНК с укороченными поли (A) хвостами не разлагаются, а вместо этого хранятся и трансляционно неактивны. Эти короткохвостые мРНК активируются цитоплазматическим полиаденилированием после оплодотворения во время активации яйца.
У животных поли (A) рибонуклеаза (PARN ) может связываться с 5'-шапочкой и удалите нуклеотиды из поли (А) хвоста. Уровень доступа к 5'-кэпу и поли (А) хвосту важен для контроля того, как скоро мРНК деградирует. PARN деаденилирует меньше, если РНК связывается факторами инициации 4E (на 5'-кэпе) и 4G (на поли (A) хвосте), поэтому трансляция снижает деаденилирование. Скорость деаденилирования также может регулироваться РНК-связывающими белками. После удаления поли (A) -хвоста комплекс декапирования удаляет 5'-кэп, что приводит к деградации РНК. Несколько других белков участвуют в деаденилировании в почкующихся дрожжах и клетках человека, в первую очередь в комплексе CCR4-Not.
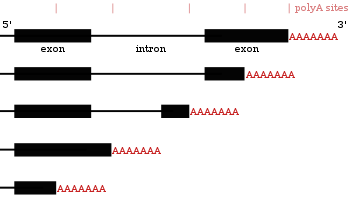 Результаты использования другого полиаденилирования сайты в одном и том же гене
Результаты использования другого полиаденилирования сайты в одном и том же гене Многие гены, кодирующие белок, имеют более одного сайта полиаденилирования, поэтому ген может кодировать несколько мРНК, которые различаются на своем 3'-конце. Поскольку альтернативное полиаденилирование изменяет длину 3'-нетранслируемой области, оно может изменять, какие сайты связывания для микроРНК содержит 3'-нетранслируемая область. МикроРНК имеют тенденцию подавлять трансляцию и способствовать деградации мРНК, с которыми они связываются, хотя есть примеры микроРНК, которые стабилизируют транскрипты. Альтернативное полиаденилирование также может укорачивать кодирующую область, тем самым создавая код мРНК для другого белка, но это гораздо менее распространено, чем просто укорачивание 3'-нетранслируемой области.
Выбор сайта поли (A) может быть зависит от внеклеточных стимулов и зависит от экспрессии белков, участвующих в полиаденилировании. Например, экспрессия CstF-64, субъединицы фактора стимуляции расщепления (CstF), увеличивается в макрофагах в ответ на липополисахариды (группа бактериальных соединений, вызывающих иммунный ответ). Это приводит к отбору слабых поли (A) сайтов и, следовательно, более коротких транскриптов. Это удаляет регуляторные элементы в 3'-нетранслируемых областях мРНК для продуктов, связанных с защитой, таких как лизоцим и TNF-α. Эти мРНК имеют более длительный период полураспада и продуцируют больше этих белков. РНК-связывающие белки, не входящие в механизм полиаденилирования, также могут влиять на использование сайта полиаденилирования, как и метилирование ДНК рядом с сигналом полиаденилирования.
Полиаденилирование происходит в цитозоле некоторых типов клеток животных, а именно в зародышевой линии, во время раннего эмбриогенеза и в пост- синаптических участках нерва. ячейки. Это удлиняет поли (А) хвост мРНК с укороченным поли (А) хвостом, так что мРНК будет транслироваться. Эти укороченные поли (A) хвосты часто состоят из менее 20 нуклеотидов и удлиняются примерно до 80–150 нуклеотидов.
У ранних эмбрионов мыши цитоплазматическое полиаденилирование материнских РНК из яйцеклетки позволяет клетке выжить и расти, даже если транскрипция не начинается до середины 2-х клеточной стадии (4-х клеточная стадия у человека). В головном мозге цитоплазматическое полиаденилирование активно во время обучения и может играть роль в долгосрочной потенциации, которая является усилением передачи сигнала от нервной клетки к другой в ответ на нервные импульсы и имеет важное значение для обучение и формирование памяти.
Цитоплазматический полиаденилирование требует РНК-связывающих белков CPSF и CPEB и может включать другие РНК-связывающие белки, такие как Pumilio. В зависимости от типа клетки полимераза может быть полиаденилат-полимеразой (PAP) того же типа, что и в ядерном процессе, или цитоплазматической полимеразой GLD-2.
Для многих некодирующих РНК, включая тРНК, рРНК, мяРНК и мяРНК, полиаденилирование способ маркировки РНК для деградации, по крайней мере, в дрожжах. Это полиаденилирование осуществляется в ядре комплексом TRAMP, который поддерживает хвост длиной около 4 нуклеотидов до 3'-конца. Затем РНК расщепляется экзосомой. Поли (A) хвосты также были обнаружены на фрагментах рРНК человека, как в форме гомополимерных (только A), так и гетерполимерных (в основном A) хвостов.
 Полиаденилирование в бактериях помогает полинуклеотидфосфорилазе разрушить прошлую вторичную структуру
Полиаденилирование в бактериях помогает полинуклеотидфосфорилазе разрушить прошлую вторичную структуру У многих бактерий полиаденилироваться могут как мРНК, так и некодирующие РНК. Этот поли (A) хвост способствует разложению деградосомой, которая содержит два фермента, разрушающих РНК: полинуклеотидфосфорилазу и РНКазу E . Полинуклеотидфосфорилаза связывается с 3'-концом РНК, и 3'-удлинение, обеспечиваемое поли (A) хвостом, позволяет ей связываться с РНК, вторичная структура в противном случае блокировала бы 3'-конец. Последовательные раунды полиаденилирования и деградации 3'-конца полинуклеотидфосфорилазой позволяют деградосоме преодолеть эти вторичные структуры. Поли (A) хвост может также привлекать РНКазы, которые разрезают РНК надвое. Эти бактериальные поли (A) хвосты имеют длину около 30 нуклеотидов.
В различных группах, таких как животные и трипаносомы, митохондрии содержат как стабилизирующие, так и дестабилизирующие поли (A)) хвосты. Дестабилизирующее полиаденилирование нацелено как на мРНК, так и на некодирующие РНК. Поли (А) хвосты в среднем имеют длину 43 нуклеотида. Стабилизирующие из них начинаются со стоп-кодона, и без них стоп-кодон (UAA) не является полным, поскольку геном кодирует только часть U или UA. Митохондрии растений обладают только дестабилизирующим полиаденилированием, а митохондрии дрожжей вообще не имеют полиаденилирования.
Хотя многие бактерии и митохондрии имеют полиаденилат-полимеразы, они также имеют другой тип полиаденилирования, осуществляемый самой полинуклеотидфосфорилазой. Этот фермент обнаружен в бактериях, митохондриях, пластидах и как составная часть экзосомы архей (в тех архей, которые имеют экзосому ). Он может синтезировать 3'-расширение, где подавляющее большинство оснований являются аденинами. Как и у бактерий, полиаденилирование полинуклеотидфосфорилазой способствует деградации РНК в пластидах и, вероятно, также в архее.
Хотя полиаденилирование наблюдается почти у всех организмов, оно не универсально. Однако широкое распространение этой модификации и тот факт, что она присутствует в организмах из всех трех доменов жизни, подразумевает, что последний универсальный общий предок всех живых организмов, считается, имел некоторую форму системы полиаденилирования. Некоторые организмы не полиаденилируют мРНК, что означает, что они утратили свои механизмы полиаденилирования в процессе эволюции. Хотя примеры эукариот, лишенных полиаденилирования, неизвестны, мРНК бактерии Mycoplasma gallisepticum и солеустойчивой археи Haloferax volcanii лишены этой модификации.
Самая древняя. ферментом полиаденилирования является полинуклеотидфосфорилаза. Этот фермент является частью бактериальной деградосомы и архейной экзосомы, двух близкородственных комплексов, которые рециклируют РНК в нуклеотиды. Этот фермент разрушает РНК, атакуя связь между 3'-крайними нуклеотидами с фосфатом, разрывая дифосфатный нуклеотид. Эта реакция обратима, поэтому фермент может также расширить РНК с помощью большего количества нуклеотидов. Гетерополимерный хвост, добавленный полинуклеотидфосфорилазой, очень богат аденином. Выбор аденина, скорее всего, является результатом более высоких концентраций АДФ, чем других нуклеотидов, в результате использования АТФ в качестве энергетической валюты, что увеличивает вероятность его включения в этот хвост в ранние формы жизни. Было высказано предположение, что участие богатых аденином хвостов в деградации РНК привело к более поздней эволюции полиаденилатных полимераз (ферментов, которые производят поли (А) хвосты без других нуклеотидов в них).
Полиаденилатные полимеразы не являются как древний. Они отдельно развились как у бактерий, так и у эукариот из CCA-добавляющего фермента, который является ферментом, завершающим 3'-концы тРНК. Его каталитический домен гомологичен другим полимеразам. Предполагается, что горизонтальный перенос бактериального фермента, добавляющего CCA, к эукариотам, позволил архейподобному ферменту, добавляющему CCA, переключить функцию на поли (A) полимеразу. Некоторые линии, такие как археи и цианобактерии, никогда не вырабатывали полиаденилат-полимеразу.
Полиаденилатные хвосты наблюдаются у нескольких РНК-вирусов, включая Грипп A, Коронавирус, Вирус мозаики люцерны и Гепатит A уток. Некоторые вирусы, такие как ВИЧ-1 и полиовирус, ингибируют поли-A-связывающий белок клетки (PABPC1 ), чтобы подчеркнуть экспрессию собственных генов над
Поли (A) -полимераза была впервые идентифицирована в 1960 году как ферментативная активность в экстрактах, сделанных из ядер клеток, которые могут полимеризовать АТФ, но не АДФ в полиаденин. Хотя эта активность идентифицирована во многих типах клеток, эта активность не имела известной функции до 1971 года, когда последовательности поли (A) были обнаружены в мРНК. Первоначально считалось, что единственной функцией этих последовательностей является защита 3'-конца РНК от нуклеаз, но позже были идентифицированы специфические роли полиаденилирования в ядерном экспорте и трансляции. Полимеразы, ответственные за полиаденилирование, были впервые очищены и охарактеризованы в 1960-х и 1970-х годах, но большое количество дополнительных белков, контролирующих этот процесс, было обнаружено только в начале 1990-х.