
Войти
| Просмотр / редактирование человека | Просмотр / редактирование мыши |
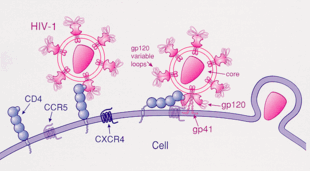 Присоединение ВИЧ к CD4 + Т-хелперной клетке: 1) вирус gp120 белок прикрепляется к CD4. 2) переменная петля gp120 присоединяется к корецептору, CCR5 или CXCR4. 3) ВИЧ проникает в клетку.
Присоединение ВИЧ к CD4 + Т-хелперной клетке: 1) вирус gp120 белок прикрепляется к CD4. 2) переменная петля gp120 присоединяется к корецептору, CCR5 или CXCR4. 3) ВИЧ проникает в клетку. CC-хемокиновый рецептор 5, также известный как CCR5 или CD195, представляет собой белок на поверхности лейкоцитов, которые участвуют в иммунной системе, поскольку они действуют как рецептор для хемокинов.
У человека ген CCR5, который кодирует белок CCR5, расположен на коротком (p) плече в положении 21 на хромосоме 3. Некоторые популяции унаследовали мутацию Delta 32, что привело к генетической делеции части гена CCR5. Гомозиготные носители этой мутации устойчивы к М-тропным штаммам инфекции ВИЧ-1.
Белок CCR5 принадлежит к семейству бета-хемокиновых рецепторов интегральных мембранных белков. Это рецептор, связанный с G-белком, который функционирует как хемокиновый рецептор в группе хемокинов CC.
Родственные CCR5 лиганды включают CCL3, CCL4 (также известный как MIP 1α и 1β соответственно) и CCL3L1. CCR5, кроме того, взаимодействует с CCL5 (хемотаксический цитокиновый белок, также известный как RANTES).
CCR5 преимущественно экспрессируется на Т-клетках, макрофагах, дендритных клетках, эозинофилах, микроглии и субпопуляции молочной железы или простаты раковые клетки. Экспрессия CCR5 избирательно индуцируется во время процесса трансформации рака и не экспрессируется в нормальных эпителиальных клетках молочной железы или предстательной железы. Приблизительно 50% рака груди человека экспрессирует CCR5, в первую очередь при тройном отрицательном раке груди. Ингибиторы CCR5 блокируют миграцию и метастазирование клеток рака груди и простаты, экспрессирующих CCR5, что позволяет предположить, что CCR5 может функционировать как новая терапевтическая мишень. Недавние исследования показывают, что CCR5 экспрессируется в подгруппе раковых клеток с характеристиками раковых стволовых клеток, которые, как известно, вызывают резистентность к терапии, и что ингибиторы CCR5 увеличивают количество клеток, убитых текущей химиотерапией. Вероятно, что CCR5 играет роль в воспалительных ответах на инфекцию, хотя его точная роль в нормальной иммунной функции неясна. Области этого белка также имеют решающее значение для связывания хемокинового лиганда, функционального ответа рецептора и активности корецептора ВИЧ.
 Последовательность первичного белка
Последовательность первичного белка ВИЧ-1 чаще всего использует хемокин рецепторы CCR5 и / или CXCR4 в качестве корецепторов для проникновения в иммунологические клетки-мишени. Эти рецепторы расположены на поверхности иммунных клеток хозяина, благодаря чему они обеспечивают способ проникновения вируса ВИЧ-1 для инфицирования клетки. Структура гликопротеина оболочки ВИЧ-1 важна для обеспечения проникновения вируса ВИЧ-1 в целевую клетку-хозяин. Структура гликопротеина оболочки состоит из двух белковых субъединиц, отщепленных от предшественника белка Gp160, кодируемого геном env ВИЧ-1: внешней субъединицы Gp120 и трансмембранной субъединицы Gp41. Эта структура гликопротеина оболочки расположена в виде шиповидной структуры, расположенной на поверхности вириона, и состоит из тримеров гетеродимеров Gp120-Gp41. Белок оболочки Gp120 является имитатором хемокинов. Хотя ему не хватает уникальной структуры хемокина, он все же способен связываться с хемокиновыми рецепторами CCR5 и CXCR4. Во время инфицирования ВИЧ-1 субъединица гликопротеина оболочки Gp120 связывается с гликопротеином CD4 и корецептором ВИЧ-1, экспрессируемым на клетке-мишени, образуя гетеротримерный комплекс. Образование этого комплекса стимулирует высвобождение фузогенного пептида, заставляя вирусную мембрану сливаться с мембраной целевой клетки-хозяина. Поскольку связывание только с CD4 может иногда приводить к отщеплению gp120, gp120 должен затем связываться с корецептором CCR5, чтобы слияние продолжалось. Тирозин-сульфированный амино-конец этого корецептора является «существенным детерминантом» связывания с гликопротеином gp120. Корецептор также распознает область V1-V2 gp120 и мостиковый лист (антипараллельный четырехцепочечный β-лист, который соединяет внутренний и внешний домены gp120). Ствол V1-V2 может влиять на «использование корецептора через свой пептидный состав, а также степенью N-связанного гликозилирования». Однако в отличие от V1-V2 петля V3 очень вариабельна и, таким образом, является наиболее важным детерминантом специфичности корецептора. Нормальные лиганды для этого рецептора, RANTES, MIP-1β и MIP-1α, способны подавлять инфекцию ВИЧ-1. in vitro. У людей, инфицированных ВИЧ, вирусы, использующие CCR5, являются преобладающими видами, выделенными на ранних стадиях вирусной инфекции, что позволяет предположить, что эти вирусы могут иметь селективное преимущество во время передачи или острой фазы заболевания. Более того, по крайней мере, половина всех инфицированных на протяжении всего периода заражения несет в себе только вирусы, использующие CCR5.
CCR5 - это первичный корецептор, используемый gp120 последовательно с CD4. Это связывание приводит к тому, что gp41, другой белковый продукт gp160, высвобождается из своей метастабильной конформации и внедряется в мембрану клетки-хозяина. Хотя это не было подтверждено, связывание gp120-CCR5 включает два важных этапа: 1) Тирозин-сульфированный амино-конец этого корецептора является «существенной детерминантой» связывания с gp120 (как указано ранее) 2) После этапа 1., должно быть реципрокное действие (синергизм, взаимосвязь) между gp120 и трансмембранными доменами CCR5.
CCR5 необходим для распространения штамма R5 вируса ВИЧ-1. Знание механизма, с помощью которого этот штамм ВИЧ-1 опосредует инфекцию, побудило исследовать разработку терапевтических вмешательств для блокирования функции CCR5. Был разработан ряд новых экспериментальных препаратов против ВИЧ, называемых антагонистами рецептора CCR5, которые препятствуют связыванию между белком оболочки Gp120 и корецептором CCR5 ВИЧ. Эти экспериментальные препараты включают PRO140 (), Vicriviroc (испытания фазы III были отменены в июле 2010 г.) (Schering Plough ), Aplaviroc ( GW-873140) (GlaxoSmithKline ) и Maraviroc (UK-427857) (Pfizer ). Маравирок был одобрен для использования FDA в августе 2007 года. Это единственный препарат, одобренный FDA для клинического применения, таким образом, он стал первым ингибитором CCR5. Проблема этого подхода состоит в том, что, хотя CCR5 является основным корецептором, с помощью которого ВИЧ заражает клетки, он не единственный такой корецептор. Возможно, что под давлением отбора ВИЧ эволюционирует и использует другой корецептор. Однако исследование вирусной устойчивости к AD101, молекулярному антагонисту CCR5, показало, что устойчивые вирусы не переключались на другой корецептор (CXCR4), но продолжали использовать CCR5: они либо связывались с альтернативными доменами CCR5. или к рецептору с более высоким сродством. Однако, поскольку существует еще один доступный корецептор, вероятно, что отсутствие гена CCR5 не делает человека невосприимчивым к вирусу; для человека было бы просто сложнее заразиться этим. Также у вируса есть доступ к CD4. В отличие от CCR5, который не требуется (что подтверждается теми, кто ведет здоровый образ жизни даже при отсутствии гена в результате мутации delta32), CD4 имеет решающее значение для системы иммунной защиты организма. Даже при отсутствии корецептора (даже CCR5) вирус все еще может проникать в клетки, если gp41 претерпит изменения (включая его цитоплазматический хвост), которые привели к независимости CD4 без необходимости CCR5 и / или CXCR4.
Экспрессия CCR5 индуцируется в эпителиальных клетках груди и простаты после трансформации. Индукция экспрессии CCR5 способствует клеточной инвазии, миграции и метастазированию. Для индукции метастазирования необходимо вернуться к месту метастазирования. Было показано, что ингибиторы CCR5, включая маравирок и леронлимаб, блокируют метастазирование в легкие клеточных линий рака молочной железы человека. В доклинических исследованиях иммунокомпетентных мышей ингибиторы CCR5 блокировали метастазирование в кости и мозг. Ингибиторы CCR5 также уменьшают инфильтрацию ассоциированных с опухолью макрофагов. Фаза 1 клинического исследования ингибитора CCR5 у пациентов с метастатическим раком толстой кишки, подвергавшихся интенсивному предварительному лечению, продемонстрировала объективный клинический ответ и снижение метастатической опухолевой нагрузки.
Повышенные уровни CCR5 являются частью воспалительная реакция на инсульт. Блокирование CCR5 с помощью маравирок (препарат, одобренный для лечения ВИЧ) может ускорить восстановление после инсульта.
В развивающемся мозге хемокины, такие как CCR5, влияют на миграцию и связь нейронов. После инсульта они, по-видимому, уменьшают количество участков соединения на нейронах, находящихся рядом с повреждением.
CCR5-Δ32 (или CCR5-D32 или CCR5 delta 32) составляет аллель CCR5.
CCR5 Δ32 представляет собой делецию из 32 пар оснований, которая вводит преждевременный стоп-кодон в локус рецептора CCR5, в результате чего рецептор становится нефункциональным. CCR5 необходим для проникновения M-тропного вируса ВИЧ-1. Лица , гомозиготные (обозначенные Δ32 / Δ32) по CCR5 Δ32, не экспрессируют функциональные рецепторы CCR5 на поверхности своих клеток и устойчивы к инфекции ВИЧ-1, несмотря на множественные воздействия высокого риска. Индивидуумы гетерозиготные (+ / Δ32) по мутантному аллелю имеют более чем 50% снижение функциональных рецепторов CCR5 на поверхности их клеток из-за димеризации между мутантными рецепторами и рецепторами дикого типа, которая мешает с транспортом CCR5 к поверхности клетки. Гетерозиготы-носители устойчивы к инфекции ВИЧ-1 по сравнению с дикими типами, а при инфицировании гетерозиготы демонстрируют сниженную вирусную нагрузку и на 2-3 года медленнее прогрессируют до СПИДа по сравнению с дикими типами.. Также было показано, что гетерозиготность по этому мутантному аллелю улучшает вирусологический ответ на антиретровирусное лечение. CCR5 Δ32 имеет частоту (гетерозиготного) аллеля 10% в Европе и частоту гомозигот 1%.
Недавние исследования показывают, что CCR5 Δ32 улучшает познавательные способности и память. В 2016 году исследователи показали, что удаление гена CCR5 у мышей значительно улучшило их память. CCR5 - мощный супрессор нейронов пластичности, обучения и памяти; Чрезмерная активация CCR5 вирусными белками может способствовать когнитивному дефициту, связанному с ВИЧ.
Аллель CCR5 Δ32 отличается недавним происхождением, неожиданно высокой частотой, и четкое географическое распространение, которые вместе предполагают, что (а) оно возникло в результате единственной мутации и (б) исторически было предметом положительного отбора.
Два исследования использовали анализ сцепления для оценки возраста делеции CCR5 Δ32, предполагая, что количество рекомбинации и мутации, наблюдаемых в геномных областях, окружающих делецию CCR5 Δ32, будет пропорционально возрасту делеции. Используя выборку из 4000 человек из 38 этнических групп, Stephens et al. подсчитали, что делеция CCR5-Δ32 произошла 700 лет назад (275-1875, 95% доверительный интервал). Другая группа, Libert et al. (1998) использовали микросателлитные мутации для оценки возраста мутации CCR5 Δ32 как 2100 лет (700–4800, 95% доверительный интервал). На основе наблюдаемых событий рекомбинации они оценили возраст мутации в 2250 лет (900-4700, 95% доверительный интервал). Третья гипотеза опирается на градиент частоты аллелей с севера на юг в Европе, который показывает, что самая высокая частота аллелей наблюдается в северных странах, а самая низкая частота аллелей - в южной Европе. Поскольку викинги исторически населяли эти страны, возможно, что аллель распространился по Европе из-за расселения викингов в 8-10 веках. Позже викинги были заменены варягами в России, что, возможно, способствовало наблюдаемому изменению частоты аллелей с востока на запад.
Первоначально ВИЧ-1 передавался от шимпанзе (Pan troglodytes) для людей в начале 1900-х годов в Юго-Восточном Камеруне, Африка, в результате контакта с инфицированной кровью и биологическими жидкостями во время разделки мяса диких животных. Однако до 1980-х годов ВИЧ-1 фактически отсутствовал в Европе. Таким образом, учитывая средний возраст примерно 1000 лет для аллеля CCR5-Δ32, можно установить, что ВИЧ-1 не оказывал давления отбора на человеческую популяцию достаточно долго, чтобы достичь нынешних частот. Следовательно, другие патогены были предложены в качестве агентов положительной селекции на CCR5 Δ32, включая бубонную чуму (Yersinia pestis ) и оспу (Variola майор ). Другие данные предполагают, что частота аллелей испытывала отрицательное давление отбора в результате патогенов, которые стали более распространенными во время римской экспансии. Идея о том, что отрицательный отбор играет роль в низкой частоте аллеля, также подтверждается экспериментами с использованием мышей с нокаутом и вирусом гриппа A, которые продемонстрировали, что присутствие рецептора CCR5 важно для эффективного ответа на патоген.
Несколько линий доказательств предполагают, что аллель CCR5 Δ32 эволюционировал только один раз. Во-первых, CCR5 Δ32 имеет относительно высокую частоту в нескольких различных европейских популяциях, но сравнительно отсутствует в популяциях азиатских, ближневосточных и американских индейцев, что позволяет предположить, что единственная мутация произошла после расхождения европейцев от их африканских предков. Во-вторых, анализ генетического сцепления показывает, что мутация происходит на гомогенном генетическом фоне, подразумевая, что мутация унаследована от общего предка. Это было продемонстрировано, показывая, что аллель CCR5 Δ32 находится в сильном неравновесном сцеплении с высокополиморфными микросателлитами. Более 95% хромосом CCR5 Δ32 также несли аллель IRI3.1-0, а 88% несли аллель IRI3.2. Напротив, микросателлитные маркеры IRI3.1-0 и IRI3.2-0 были обнаружены только в 2 или 1,5% хромосом, несущих аллель CCR5 дикого типа. Это свидетельство неравновесия по сцеплению поддерживает гипотезу о том, что большинство, если не все, аллели CCR5 Δ32 возникли в результате одного мутационного события. Наконец, аллель CCR5 Δ32 имеет уникальное географическое распределение, указывающее на единственное северное происхождение с последующей миграцией. Исследование, измеряющее частоты аллелей в 18 европейских популяциях, обнаружило градиент с севера на юг, с самыми высокими частотами аллелей в финских и мордовских популяциях (16%) и самыми низкими в Сардинии (4%). 125>
В отсутствие отбора одной мутации потребуется примерно 127 500 лет, чтобы вырасти до 10% популяционной частоты. По оценкам, основанным на генетической рекомбинации и частоте мутаций, возраст аллеля составляет от 1000 до 2000 лет. Это несоответствие является признаком положительного отбора.
Подсчитано, что ВИЧ-1 проник в популяцию людей в Африке в начале 1900-х годов; однако о симптоматических инфекциях не сообщалось до 1980-х годов. Следовательно, эпидемия ВИЧ-1 еще слишком молода, чтобы быть источником положительного отбора, который привел к тому, что частота CCR5 Δ32 с нуля до 10% за 2000 лет. Стивенс и др. (1998), предполагают, что бубонная чума (Yersinia pestis) оказывала положительное селективное давление на CCR5 Δ32. Эта гипотеза была основана на сроках и серьезности пандемии «черной смерти», в результате которой погибло 30% европейского населения всех возрастов между 1346 и 1352 годами. После «черной смерти» наблюдались менее серьезные периодические эпидемии. В отдельных городах наблюдалась высокая смертность, но общая смертность в Европе составляла всего несколько процентов. В 1655–1656 годах вторая пандемия, названная «Великая чума», унесла жизни 15-20% населения Европы. Важно отметить, что эпидемии чумы носили прерывистый характер. Бубонная чума - это зоонозное заболевание, которое поражает в первую очередь грызунов, распространяется блохами и лишь изредка заражает людей. Передача от человека человеку бубонной чумы не происходит, но может возникнуть при легочной чуме, которая поражает легкие. Только при низкой плотности грызунов инфицированные блохи вынуждены питаться альтернативными хозяевами, такими как люди, и в этих обстоятельствах может возникнуть человеческая эпидемия. Основываясь на популяционно-генетических моделях, Гальвани и Слаткин (2003) утверждают, что прерывистый характер эпидемий чумы не создавал достаточно сильной селективной силы, чтобы довести частоту аллелей CCR5 Δ32 до 10% в Европе.
Чтобы проверить В соответствии с этой гипотезой Гальвани и Слаткин (2003) смоделировали историческое давление отбора, вызванное чумой и оспой. Чума была смоделирована в соответствии с историческими данными, в то время как возрастная смертность от оспы была определена на основе возрастного распределения захоронений оспы в Йорке (Англия) между 1770 и 1812 годами. Оспа преимущественно поражает молодых, до репродуктивных членов населения, поскольку они являются единственными лица, не прошедшие вакцинацию, или умершие от перенесенной инфекции. Поскольку оспа преимущественно убивает представителей популяции до репродуктивного возраста, она создает более сильное избирательное давление, чем чума. В отличие от чумы, оспа не имеет животного резервуара и передается только от человека к человеку. Авторы подсчитали, что если бы чума отбиралась по CCR5 Δ32, частота аллеля все равно была бы менее 1%, в то время как оспа оказывала селективную силу, достаточную для достижения 10%.
Гипотеза о том, что натуральная оспа оказывала положительный отбор на CCR5 Δ32, также является биологически правдоподобной, поскольку поксвирусы, такие как ВИЧ, проникают в лейкоциты с помощью хемокиновых рецепторов. Напротив, Yersinia pestis - это бактерия с совершенно другой биологией.
Хотя европейцы являются единственной группой, в которой есть субпопуляции с высокой частотой CCR5 Δ32, они не единственная популяция, которая подверглась селекции с помощью натуральной оспы, которая имела всемирное распространение до того, как была объявлена искорененной в 1980 году. Самые ранние безошибочные описания оспы появляются в 5 веке нашей эры в Китае, в 7 веке нашей эры в Индии и Средиземноморье и в 10 веке нашей эры в Юго-Западной Азии. Напротив, мутация CCR5 Δ32 обнаруживается только в популяциях Европы, Западной Азии и Северной Африки. Аномально высокая частота CCR5 Δ32 в этих популяциях, по-видимому, требует как уникального происхождения в Северной Европе, так и последующего отбора оспой.
CCR5 Δ32 могут быть полезны для хозяина при одних инфекциях (например, ВИЧ-1, возможно, оспа), но пагубны при других (например, клещевой энцефалит, Западный Нил). вирус). Будет ли функция CCR5 полезна или вредна в контексте данной инфекции, зависит от сложного взаимодействия между иммунной системой и патогеном.
В целом исследования показывают, что мутация CCR5 Δ32 может играть пагубную роль в постинфекционных воспалительных процессах, которые могут повреждать ткань и создавать дополнительную патологию. Лучшее доказательство этой предполагаемой антагонистической плейотропии обнаружено при инфекциях флавивирусом. Как правило, многие вирусные инфекции протекают бессимптомно или вызывают лишь легкие симптомы у подавляющего большинства населения. Однако у некоторых незадачливых людей протекает особенно разрушительное клиническое течение, которое иначе необъяснимо, но, по-видимому, является генетически опосредованным. Было обнаружено, что пациенты, гомозиготные по CCR5 Δ32, имеют более высокий риск нейроинвазивной формы клещевого энцефалита (флавивирус). Кроме того, функциональный CCR5 может потребоваться для предотвращения симптоматического заболевания после инфицирования вирусом Западного Нила, другим флавивирусом; CCR5 Δ32 был связан с ранним развитием симптомов и более выраженными клиническими проявлениями после инфицирования вирусом Западного Нила.
Это открытие у людей подтвердило ранее наблюдаемый эксперимент на животной модели гомозиготности CCR5 Δ32. После инфицирования вирусом Западного Нила у мышей CCR5 Δ32 были заметно увеличены вирусные титры в центральной нервной системе и повышена смертность по сравнению с мышами дикого типа, что позволяет предположить, что экспрессия CCR5 необходима для создания сильной защиты хозяина от вируса Западного Нила..
Генетический подход с использованием внутриклеточных тел, которые блокируют экспрессию CCR5, был предложен в качестве лечения ВИЧ-1 инфицированных людей. Когда Т-клетки модифицировались так, что они больше не экспрессируют CCR5, были смешаны с немодифицированными Т-клетками, экспрессирующими CCR5, а затем заражены инфекцией ВИЧ-1, модифицированные Т-клетки, которые не экспрессируют CCR5, в конечном итоге захватили культуру, как ВИЧ-1. убивает немодифицированные Т-клетки. Этот же метод можно использовать in vivo для создания пула устойчивых к вирусу клеток у инфицированных людей.
Эта гипотеза была проверена на пациенте со СПИДом, у которого также развился миелоидный лейкоз, и который проходил лечение с химиотерапией для подавления рака. Затем трансплантат костного мозга, содержащий стволовые клетки от подходящего донора, использовали для восстановления иммунной системы. Однако трансплантат был выполнен от донора с 2 копиями гена мутации CCR5-Δ32. Через 600 дней пациент был здоров и имел неопределяемые уровни ВИЧ в крови и исследованных тканях мозга и прямой кишки. Перед трансплантацией также были обнаружены низкие уровни ВИЧ X4, который не использует рецептор CCR5. Однако после трансплантации этот тип ВИЧ также не был обнаружен. Однако этот результат согласуется с наблюдением, что клетки, экспрессирующие вариантный белок CCR5-Δ32, лишены рецепторов CCR5 и CXCR4 на своей поверхности, что обеспечивает устойчивость к широкому спектру вариантов ВИЧ, включая HIVX4. По прошествии более шести лет у пациента сохранилась устойчивость к ВИЧ и было объявлено, что ВИЧ-инфекция вылечена.
В 2009 году началось включение ВИЧ-положительных пациентов в клинические испытания, в которых клетки пациентов были генетически модифицирован с помощью нуклеазы цинкового пальца, чтобы нести признак CCR5-Δ32, а затем повторно введен в организм в качестве потенциального средства лечения ВИЧ. Результаты, опубликованные в 2014 году, были многообещающими.
Вдохновленный первым человеком, когда-либо излечившимся от ВИЧ, пациентом из Берлина, StemCyte начал сотрудничество с банками пуповинной крови по всему миру, чтобы систематически проверять образцы пуповинной крови на наличие мутации CCR5, начиная с 2011 года.
В ноябре 2018 года Цзянькуй Хэ объявил, что он отредактировал два человеческих эмбриона, чтобы попытаться отключить ген CCR5, который кодирует рецептор, который ВИЧ использует для проникновения в клетки. Он сказал, что девочки-близнецы, Лулу и Нана, родились несколькими неделями ранее, и что девочки все еще несут функциональные копии CCR5 вместе с отключенным CCR5 (мозаицизм ), следовательно, все еще уязвимы для ВИЧ. Работа была широко осуждена как неэтичная, опасная и преждевременная.




