
Войти
An интрон (для внутригенной области) представляет собой любую нуклеотидную последовательность в гене, которая удаляется сплайсингом РНК во время созревания конечный продукт РНК. Другими словами, интроны представляют собой некодирующие области транскрипта РНК или кодирующей его ДНК, которые удаляются путем сплайсинга перед трансляцией. Слово интрон происходит от термина внутригенная область, то есть область внутри гена. Термин интрон относится как к последовательности ДНК в гене, так и к соответствующей последовательности в транскриптах РНК . Последовательности, которые объединяются в конечную зрелую РНК после сплайсинга РНК, представляют собой экзоны.
Интроны обнаруживаются в генах большинства организмов и многих вирусов и могут быть локализованы в широком диапазоне генов, включая те, которые генерируют белки, рибосомная РНК (рРНК) и транспортная РНК (тРНК). Когда белки генерируются из генов, содержащих интрон, сплайсинг РНК происходит как часть пути обработки РНК, которая следует за транскрипцией и предшествует трансляции.
Интроны были впервые обнаружены в генах, кодирующих белок аденовируса, и впоследствии были идентифицированы в генах, кодирующих гены транспортной РНК и рибосомной РНК. В настоящее время известно, что интроны встречаются в большом количестве генов организмов и вирусов во всех биологических царствах.
Тот факт, что гены были расщеплены или прерваны интронами, был независимо открыт в 1977 году Филипом Алленом Шарпом и Ричардом Дж. Робертсом для который они разделили Нобелевской премией по физиологии и медицине в 1993 году. Термин интрон был введен американским биохимиком Уолтером Гилбертом :
«Понятие цистрона [то есть ген]... должен быть заменен единицей транскрипции, содержащей области, которые будут потеряны из зрелого мессенджера - я предлагаю, мы называем интронами (для внутригенных областей) - чередующиеся с участками, которые будут экспрессироваться - экзоны ". (Gilbert 1978)
Термин интрон также относится к интрацистрону, то есть к дополнительному фрагменту ДНК, который возникает внутри цистрона.
Хотя интроны иногда называют промежуточными последовательностями, термин «промежуточная последовательность» может относиться к любое из нескольких семейств внутренних последовательностей нуклеиновых кислот, которые не присутствуют в конечном продукте гена, включая интеины, нетранслируемые последовательности (UTR ) и нуклеотиды, удаляемые, помимо интронов.
Наблюдается, что частота интронов в разных геномах широко варьируется по спектру биологических организмов. Например, интроны чрезвычайно распространены в ядерном геноме челюстных позвоночных (например, людей и мышей), где гены, кодирующие белок, почти всегда содержат несколько интронов, в то время как интроны редко встречаются в ядерных генах некоторых эукариотических микроорганизмов, например дрожжи пекарские / пивные (Saccharomyces cerevisiae). Напротив, митохондриальные геномы позвоночных полностью лишены интронов, тогда как геномы эукариотических микроорганизмов могут содержать много интронов.
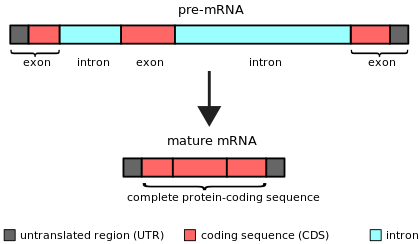 Простая иллюстрация несплайсированного предшественника мРНК с двумя интронами и тремя экзонами (вверху). После удаления интронов путем сплайсинга последовательность зрелой мРНК готова к трансляции (внизу).
Простая иллюстрация несплайсированного предшественника мРНК с двумя интронами и тремя экзонами (вверху). После удаления интронов путем сплайсинга последовательность зрелой мРНК готова к трансляции (внизу). Особенно крайним случаем является ген dhc7 Drosophila, содержащий интрон размером ≥3,6 мегабаз (МБ), для транскрипции которого требуется примерно три дня.. С другой стороны, недавнее исследование предполагает, что самая короткая известная длина интрона эукариот составляет 30 пар оснований (п.н.), принадлежащих гену MST1L человека.
Сплайсинг всех интрон-содержащих РНК молекулы внешне похожи, как описано выше. Однако различные типы интронов были идентифицированы посредством изучения структуры интронов с помощью анализа последовательности ДНК, а также генетического и биохимического анализа реакций сплайсинга РНК.
Идентифицировано по крайней мере четыре различных класса интронов:
интроны группы III, как предполагается, являются пятым семейством, но мало что известно о биохимическом аппарате, который опосредует их сплайсинг. Они, по-видимому, связаны с интронами группы II и, возможно, с интронами сплайсосом.
Интроны ядерной пре-мРНК (сплайсосомные интроны) характеризуются специфическими последовательностями интронов, расположенными на границах между интроны и экзоны. Эти последовательности распознаются молекулами сплайсосомной РНК, когда инициируются реакции сплайсинга. Кроме того, они содержат точку ветвления, конкретную нуклеотидную последовательность около 3'-конца интрона, которая становится ковалентно связанной с 5'-концом интрона в процессе сплайсинга, образуя разветвленный (лариатный) интрон. Помимо этих трех коротких консервативных элементов, интронные последовательности ядерной пре-мРНК очень вариабельны. Интроны ядерной пре-мРНК часто намного длиннее, чем окружающие их экзоны.
Интроны переносящей РНК, удаление которых зависит от белков, находятся в определенном месте внутри петли антикодона несплайсированных предшественников тРНК и удаляются эндонуклеазой сплайсинга тРНК. Затем экзоны связываются вместе вторым белком, лигазой сплайсинга тРНК. Обратите внимание, что самосплайсирующие интроны также иногда встречаются в генах тРНК.
Интроны группы I и группы II обнаруживаются в генах, кодирующих белки (матричная РНК ), переносят РНК и рибосомную РНК в очень широкий круг живых организмов. После транскрипции в РНК интроны группы I и группы II также осуществляют обширные внутренние взаимодействия, которые позволяют их складывать в конкретную сложную трехмерную архитектуру. Эта сложная архитектура позволяет некоторым интронам группы I и группы II самосплайсироваться, то есть содержащая интрон молекула РНК может перестроить свою собственную ковалентную структуру, чтобы точно удалить интрон и связать экзоны вместе в правильном порядке. В некоторых случаях определенные связывающие интрон белки участвуют в сплайсинге, действуя таким образом, что они помогают интрону складываться в трехмерную структуру, которая необходима для активности самосплайсинга. Интроны группы I и группы II отличаются разными наборами внутренних консервативных последовательностей и складчатых структур, а также тем фактом, что при сплайсинге молекул РНК, содержащих интроны группы II, образуются разветвленные интроны (как интроны сплайсосомных РНК), в то время как интроны группы I используют не -кодированный нуклеотид гуанозина (обычно GTP), чтобы инициировать сплайсинг, добавляя его к 5'-концу вырезанного интрона.
Хотя интроны не кодируют белковые продукты, они являются неотъемлемой частью регуляции экспрессии генов. Некоторые интроны сами кодируют функциональные РНК посредством дальнейшей обработки после сплайсинга с образованием некодирующих молекул РНК. Альтернативный сплайсинг широко используется для генерации множества белков из одного гена. Кроме того, некоторые интроны играют важную роль в широком диапазоне функций регуляции экспрессии генов, таких как нонсенс-опосредованный распад и экспорт мРНК.
Биологическое происхождение интронов неясно. После первоначального открытия интронов в генах, кодирующих белок эукариотического ядра, возникли серьезные споры о том, унаследованы ли интроны в современных организмах от общего древнего предка (так называемая гипотеза раннего интрона) или же они появились в гены сравнительно недавно в эволюционном процессе (так называемая гипотеза интронно-позднего периода). Другая теория состоит в том, что сплайсосома и структура интрон-экзон генов являются пережитком мира РНК (гипотеза «сначала интроны»). До сих пор ведутся серьезные споры о том, насколько верна какая из этих гипотез. В настоящее время широко распространено мнение о том, что интроны возникли внутри эукариотической линии как эгоистичные элементы.
Ранние исследования геномных последовательностей ДНК от широкого круга организмов показывают, что интрон-экзонная структура гомологичных генов у разных организмов может различаться. широко. Более поздние исследования полных эукариотических геномов показали, что длина и плотность (интроны / ген) интронов значительно различаются между родственными видами. Например, в то время как геном человека содержит в среднем 8,4 интронов / ген (139 418 в геноме), одноклеточный гриб Encephalitozoon cuniculi содержит только 0,0075 интронов / ген (15 интронов в геноме). Поскольку эукариоты произошли от общего предка (общее происхождение ), в течение эволюционного времени должно было происходить значительное увеличение или потеря интронов. Считается, что этот процесс подлежит отбору с тенденцией к увеличению интронов у более крупных видов из-за их меньшего размера популяции и наоборот у более мелких (особенно одноклеточных) видов. Биологические факторы также влияют на то, какие гены в геноме теряют или накапливают интроны.
Альтернативное сплайсинг экзонов внутри гена после вырезания интрона способствует большей вариабельности белковых последовательностей, транслируемых из одного гена, что позволяет использовать несколько родственных белков. генерируется из одного гена и одного транскрипта мРНК-предшественника. Контроль альтернативного сплайсинга РНК осуществляется сложной сетью сигнальных молекул, которые отвечают на широкий спектр внутриклеточных и внеклеточных сигналов.
Интроны содержат несколько коротких последовательностей, которые важны для эффективного сплайсинга, такие как акцепторные и донорные сайты на обоих концах интрона, а также сайт точки ветвления, которые необходимы для правильного сплайсинга сплайсосомой . Известно, что некоторые интроны усиливают экспрессию гена, в котором они содержатся, с помощью процесса, известного как интрон-опосредованное усиление (IME).
Активно транскрибируемые области ДНК часто образуют R-петли, которые уязвимы для повреждения ДНК. В высокоэкспрессируемых генах дрожжей интроны ингибируют образование R-петли и возникновение повреждений ДНК. Полногеномный анализ как у дрожжей, так и у людей показал, что гены, содержащие интроны, имеют пониженные уровни R-петли и снижение повреждений ДНК по сравнению с генами без интронов с аналогичной экспрессией. Вставка интрона в ген, подверженный R-петле, также может подавлять образование R-петли и рекомбинацию. Bonnet et al. (2017) предположили, что функция интронов в поддержании генетической стабильности может объяснить их эволюционное поддержание в определенных местах, особенно в сильно экспрессируемых генах.
Физическое присутствие интронов способствует устойчивости клеток к голоданию за счет усиленной интронами репрессии генов рибосомных белков в путях определения питательных веществ.
Интроны могут теряться или приобретаться с течением времени, как показывают многие сравнительные исследования ортологичных генов. Последующий анализ выявил тысячи примеров событий потери и усиления интронов, и было высказано предположение, что появление эукариот или начальные стадии эволюции эукариот связаны с вторжением интронов. Были идентифицированы два определяющих механизма потери интрона, опосредованная обратной транскриптазой потеря интрона (RTMIL) и геномные делеции, и известно, что они происходят. Однако окончательные механизмы усиления интронов остаются неуловимыми и спорными. На данный момент описано по крайней мере семь механизмов увеличения интрона: транспозиция интрона, вставка транспозона, тандемная геномная дупликация, перенос интрона, усиление интрона во время восстановления двухцепочечного разрыва (DSBR), вставка интрона группы II и интронизация. Теоретически было бы проще всего установить происхождение недавно приобретенных интронов из-за отсутствия мутаций, вызванных хозяином, но даже недавно полученные интроны не возникли ни в одном из вышеупомянутых механизмов. Таким образом, эти открытия поднимают вопрос о том, не могут ли предложенные механизмы усиления интронов описать механистическое происхождение многих новых интронов, потому что они не являются точными механизмами усиления интронов, или есть ли другие, еще не обнаруженные процессы, порождающие новые интроны.
При транспозиции интронов, наиболее часто предполагаемом механизме усиления интронов, сплайсированный интрон, как полагают, выполняет обратное сплайсинг либо в свою собственную мРНК, либо в другую мРНК в ранее не имеющем интрона положении. Эта содержащая интрон мРНК затем подвергается обратной транскрипции, и полученная содержащая интрон кДНК может затем вызвать усиление интрона посредством полной или частичной рекомбинации с исходным геномным локусом. Вставки транспозонов также могут приводить к созданию интронов. Такая вставка может интронизировать транспозон без нарушения кодирующей последовательности, когда транспозон вставляется в последовательность AGGT, что приводит к дупликации этой последовательности на каждой стороне транспозона. Пока не понятно, почему эти элементы сращиваются случайно или в результате какого-либо предпочтительного действия транспозона. При тандемной геномной дупликации из-за сходства между консенсусными донорскими и акцепторными сайтами сплайсинга, которые оба очень похожи на AGGT, тандемная геномная дупликация экзонного сегмента, несущего последовательность AGGT, генерирует два потенциальных сайта сплайсинга. При распознавании сплайсосомой последовательность между исходной и дублированной AGGT будет сплайсирована, что приведет к созданию интрона без изменения кодирующей последовательности гена. Восстановление двухцепочечных разрывов посредством негомологичного соединения концов было недавно идентифицировано как источник увеличения интронов, когда исследователи определили короткие прямые повторы, фланкирующие 43% полученных интронов у дафний. Эти числа необходимо сравнивать с числом консервативных интронов, фланкированных повторами в других организмах, однако для статистической значимости. Для вставки интрона группы II было высказано предположение, что ретросомизация интрона группы II в ядерный ген может вызвать недавнее усиление сплайсосомного интрона.
Предполагается, что перенос интрона приводит к увеличению интрона, когда паралог или псевдоген приобретает интрон и затем переносит этот интрон посредством рекомбинации в место, где отсутствует интрон, в его сестринском паралоге. Интронизация - это процесс, при котором мутации создают новые интроны из ранее существовавшей экзонной последовательности. Таким образом, в отличие от других предложенных механизмов увеличения интрона, этот механизм не требует вставки или генерации ДНК для создания нового интрона.
Единственным гипотетическим механизмом недавнего увеличения интрона, не имеющим прямых доказательств, является механизм группы II. вставка интрона, продемонстрированная in vivo, устраняет экспрессию генов. Поэтому интроны группы II, вероятно, являются предполагаемыми предками сплайсосомных интронов, действуя как сайт-специфичные ретроэлементы, и больше не отвечают за усиление интронов. Тандемная геномная дупликация - единственный предложенный механизм, подтверждающий экспериментальные данные in vivo: короткая внутригенная тандемная дупликация может вставлять новый интрон в ген, кодирующий белок, оставляя соответствующую пептидную последовательность неизменной. Этот механизм также имеет обширные косвенные доказательства, подтверждающие идею о том, что тандемная геномная дупликация является превалирующим механизмом увеличения интронов. Тестирование других предложенных механизмов in vivo, в частности, увеличения интронов во время DSBR, переноса интронов и интронизации, возможно, хотя эти механизмы должны быть продемонстрированы in vivo, чтобы закрепить их как действительные механизмы увеличения интронов. Дальнейший геномный анализ, особенно когда он выполняется на уровне популяции, может затем количественно оценить относительный вклад каждого механизма, возможно, выявить видоспецифичные отклонения, которые могут пролить свет на различные скорости увеличения интронов у разных видов.
Структура:
Сплайсинг:
Функция
Другое:
| Найдите intron в Wiktionary, бесплатном словаре. |