
Войти
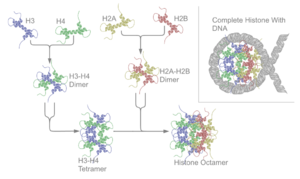 Схематическое изображение сборки ядер гистонов в нуклеосому.
Схематическое изображение сборки ядер гистонов в нуклеосому. В биологии, гистоны представляют собой высоко основные белки, обнаруженные в ядрах эукариотических клеток., которые упаковывают и упорядочивают ДНК в структурные единицы, называемые нуклеосомами. Гистоны содержат большое количество лизина и аргинина. Гистоны - главные белковые компоненты хроматина, действующие как катушки, вокруг которых наматывается ДНК, и играющие роль в регуляции генов. Без гистонов развернутая ДНК в хромосомах была бы очень длинной (отношение длины к ширине более 10 миллионов к 1 в ДНК человека). Например, каждая диплоидная клетка человека (содержащая 23 пары хромосом) имеет около 1,8 метра ДНК; ранана гистонах, диплоидная клетка имеет около 90 микрометров (0,09 мм) хроматина. Когда диплоидные клетки заменяются и конденсируются во время митоза, в результате получается около 120 микрометров хромосом.
| Ядро гистона H2A / H2B / H3 / H4 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
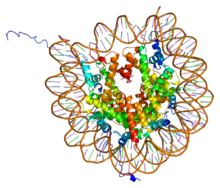 PDB рендеринг комплекса между ядерной частью нуклеосом (h3, h4, div class="ht"a, div class="ht"b) и фрагмент ДНК длиной 146 п.н. на основе 1aoi. PDB рендеринг комплекса между ядерной частью нуклеосом (h3, h4, div class="ht"a, div class="ht"b) и фрагмент ДНК длиной 146 п.н. на основе 1aoi. | |||||||||||
| Идентификаторы | |||||||||||
| Символ | Гистон | ||||||||||
| Pfam | PF00125 | ||||||||||
| Pfam клан | CL0012 | ||||||||||
| Int erPro | IPR007125 | ||||||||||
| SCOPe | 1hio / SUPFAM | ||||||||||
| |||||||||||
| линкерный гистон H1 и семейство H5 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 PDB рендеринг HIST1H1B на основе 1ghc. PDB рендеринг HIST1H1B на основе 1ghc. | |||||||||||
| Идентификаторы | |||||||||||
| Символ | Linker_histone | ||||||||||
| Pfam | PF00538 | ||||||||||
| InterPro | IPR005818 | ||||||||||
| SMART | SM00526 | ||||||||||
| SCOPe | 1hst / SUPFAM | ||||||||||
| |||||||||||
Существуют пять основных семейств гистонов: H1 / H5, H2A, H2B, H3 и H4. Гистоны H2A, H2B, H3 и H4 известны как гистоны ядра, а гистоны H1 / H5известны как гистоны-линкеры.
Все коровые гистоны существуют в виде димеров, которые схожи в том, что все они обладают складчатым доменом гистонов: три альфа-спирали, связанные двумя петлями. Именно спиральная структура позволяет взаимодействовать между отдельными димерами, особенно в стиле стиля-хвост (также называемый мотивом рукопожатия). Полученное четыре различных димера объединяет образование одного октамерного ядра нуклеосомы, размером примерно 63 углеродрем ( соленоид (ДНК) -подобная частица). Около 146 пар оснований (п.н.) ДНК оборачиваются вокруг этой частицы ядра 1,65 раза в виде левого суперспирального поворота, давая частицу размером около 100 кругрем. Линкерный гистон H1 связывает нуклеосому на участках входа и выхода ДНК, таким образом фиксируя ДНК на месте и образование структуры более высокого порядка. Самым основным из таких образований является волокно или бусинки длиной 10 нм на нити. Это включает обертывание ДНКвокруг нуклеосом примерно с 50 парами оснований ДНК, разделяющих каждую пару нуклеосом (также называемых линкером ДНК ). Структуры более высокого порядка включают 30-нм волокно (образующее неправильное зигзаг) и 100-нм волокно, это структуры, обнаруженные в нормальных клетках. Во время митоза и мейоза конденсированные хромосомы собираются посредством взаимодействия между нуклеосомами и другими регуляторными белками.
Гистоны подразделяются наканонические репликационно-зависимые гистоны, которые экспрессируются во время S-фазы клеточного цикла, и независимые от репликации варианты гистонов, экспрессируется в течение всего клеточного цикла. У животных гены, кодирующие канонические гистоны, обычно сгруппированы вдоль хромосомы, лишены интронов и используют стержневой петли на 3 'конце вместо polyA-хвоста. Гены, кодирующие варианты гистонов, обычно не кластеризованы, имеютинтроны, а их мРНК регулируются полиА-хвостами. Сложные многоклеточные организмы обычно большее количество вариантов гистонов, множество различных функций. В последнее время накапливаются данные роли различных вариантов гистонов. Варианты гистонов от разных организмов, их классификацию и особенности можно найти в базе данных "HistoneDB 2.0 - Варианты".
Ниже представлен список человека гистоновых белков:
| Суперсемейство | Семейство | Подсемейство | Члены |
|---|---|---|---|
| Линкер | H1 | H1F | H1F0, H1FNT, H1FOO, H1FX |
| H1H1 | HIST1H1A, HIST1H1B, HIST1H1C, HIST1H1D, HIST1H1E, HIST1H1T | ||
| Core | H2A | H2AF | H2AFB1, H2AFB2, H2AFB3, H2AFJ, H2AFV, H2AFX, H2AFY, H2AFY2, H2AFZ |
| H2A1 | HIST1H2AA, HIST1H2AB, HIST1H2AC, HIST1H2AD,HIST1H2AE, <176AG1HIST1HIST1HIST1HIST1HIST1HIST1Hist, HIST1H2AI, HIST1H2AJ, HIST1H2AK, HIST1H2AL, HIST331H2AM | ||
| H2A2 <6>HIST2H2AA3>HIST2H2AC | |||
| H2B | H2BF | H2BFM, H2BFS, H2BFWT | |
| H2B1 | HIST1H2BA, HIST1H2BB, HIST1H2BC, HIST1H2BD, HI ST1H2BE, HIST1H2BF, HIST1H2BG, HIST1H2BH, HIST1H2BI, HIST1H2BJ, <1H2BKK 359>, HIST1H2BL, HIST1H2BM, HIST1H2BN, HIST1H2BO | ||
| H2B2 | HIST2H2BE | ||
| H3 | H3A1 | ||
| H3 | H3A1, HIST1H3B, HIST1H3C 387>HIST1H3D, HIST1H3E, HIST1H3F, HIST1H3G, HIST1H3H, HIST1H3I, HIST1H3J | ||
| H3A2 | HIST2H3C | ||
| H3A3 | HIST3H3 | ||
| H4 | H41 | HIST1H4A, <118BA>HIST1H4A, <118BA>HIST1H4A, <118BA>359>, <118B>HIST 359>, HIST1H4C, HIST1H4C, HIST 359>, HIST1H4E, HIST1H4F, HIST1H4G, HIST1H4H, HIST1H4I, HIST1H4J, HIST1H4K, HIST1H4L | |
| H44 | HIST4H4 |
 Этапы сборки нуклеосомы
Этапы сборки нуклеосомы Ядро нуклеосомы состоят из двух H2A- Димеры H2B и тетрамер H3-H4, образующие две почти симметричные половина п о симметричные и третичной структуры (C2 ; одна макромолекула является зеркальным отображением другой). Димеры H2A-H2B и тетрамер H3-H4 также демонстрируют псевдодиадную симметрию. Четыре «ядерных» гистона (H2A, H2B, H3 и H4) относительно схожи по структуре и высоко консервативны в ходе эволюции, все они имеют мотив «виток спирали поворот спирали». (ДНК-связывающий белковый мотив, распознающий конкретную последовательность ДНК). У них также есть характерная черта длинных «хвостов» в одной структуре аминокислоты - это местопосттрансляционной модификации (см. Ниже).
Архейский гистон содержит только H3-H4-подобная димерная структура, состоящая из той же белка. Такие димерные структуры могут складывать в суперспираль («супернуклеосому»), на которой ДНК наматывается подобные катушкам нуклеосом. Только некоторые гистоны архей имеют хвосты.
Было высказано предположение, что гистоновые белки эволюционно связаны со спиральной расширенного ААА + АТФазного домена, С-доменом и с N-концевым доменомузнавания субстрата Clp / Белки Hsp100. Несмотря на различия в их топологии, эти три складки имеют гомологичный мотив спираль-нить-спираль (HSH).
Используя метод электронного парамагнитного резонанса спин-мечения, британские исследователи измерили расстояния между катушками, вокруг которых эукариотические клетки наматывают свою ДНК. Они определили диапазон расстояний от 59 до 70 Å.
Всего гистоны взаимодействуют с ДНК пяти типов:
Высокоосновная природа гистонов, кроме облегчения взаимодействия ДНК-гистонов, их растворимости в воде.
Гистоны подвергаются посттрансляционной модификации ферментами в первую очередь на их N-концевых хвостах, но также и в их глобулярных доменах. Такие модификации включают метилирование, цитруллинирование, ацетилирование, фосфорилирование, SUMOylation, убиквитинирование и АДФ-рибозилирование. Это влияет на их функцию генной регуляции.
В общем, имеют гены, которые активны, меньше связанного гистона, в то время как неактивные гены сильно связаны с гистонами во время интерфазы. Также представляется, что структура гистонов была эволюционно консервативной, поскольку любые вредные мутации былибы сильно дезадаптивными. Все гистоны имеют высоко положительно заряженный N-конец с множеством остатков лизина и аргинина.
Гистоны были обнаружены в 1884 году Альбрехтом Косселем. Слово «гистон» датируется концом 19 века и происходит от немецкого слова «Histon», само слово неопределенного происхождения - возможно, от греческого histanai или histos.
В начале 1960-х годов, до того, как были известны типы гистонов, известно и дотого, как стало, что гистоны высоко консервативны среди таксономически разнообразных организмов, Джеймс Ф. Боннер и его сотрудники начали их изучение белки, которые, как известно, связаны с ДНК в ядрах высших организмов. Боннер и его научный сотрудник Ру Чжи К. Хуанг показывает, что хроматин не поддерживает транскрибцию РНК в пробирке, но если гистоны извлекаются из хроматина, то РНК может быть транскрибирована из оставшейся ДНК. Их статья стала классикой цитирования. ПолТ'со и Джеймс Боннер созвали Всемирный конгресс по химии и биологии гистонов в 1964 году, на котором стало ясно, что нет единого мнения о количестве видов гистонов и что никто не знает, как они будут сравнивать, если они изолированы от разных организмов. Затем и его сотрудники разработали методы разделения одного и того же гистона из разных организмов в сотрудничестве с Эми Смитом. из Калифорнийского университета в Лос-Анджелесе. Например, они представляют собой последовательностьгистона IV очень консервативна между горохом и тимусом теленка. Однако работа над биохимическими характеристиками отдельных гистонов не взаимодействует друг с другом или с ДНК.
Также в 1960-х годах Винсент Олфри и Альфред Мирски предположил, что ацетилирование и метилирование гистонов могло обеспечить механизм транскрипции, но не имел доступа к подробному анализу, который более поздние контролеры смогли провести, как такое регулирование может быть осуществлено.ген-специфичный. До начала 1990-х годов гистоны отвергались большинством как инертный упаковочный материал для ядерной ДНК эукариот - точка зрения, частично основанная на моделях Марка Пташне и других, которые считали, что транскрипция активирует белок-ДНК и белок-белковые взаимодействия в основном с голыми матрицами ДНК, как в случае с бактериями.
В течение 1980-х Яхли Лорч и Роджер Корнберг показал, что нуклеосома на коровом промоторе предотвращает инициациютранскрипции in vitro, а Майкл Грюнштейн, что гистоны репрессируют транскрипцию в vivo, что привело к идее о нуклеосоме как о репрессоре общего гена. Считается, что облегчение репрессии связано как с модификацией гистонов, так и с комплексов ремоделирования хроматина. Винсент Олфри и Альфред Мирски ранее предложили роль модификации гистонов в активации транскрипции, рассматриваемой как молекулярное проявление эпигенетики. Майкл Грюнштейн и Дэвид Аллис поддержки этого предложения вважности ацетилирования гистонов для транскрипции в поискжах и активности активатора транскрипции Gcn5 как гистонацетилтрансферазы.
Открытие гистона H5, по-видимому, относится к 1970-м годам, и теперь он считается изоформой гистона H1.
Корневые гистоны обнаружены в ядрах эукариотических клеток и в некоторых архей, а именно в протеоархиях и Euryarchaea, но не у бактерий. Одноклеточные водоросли, известные как динофлагелляты, ранее считанные единственными эукариотами, у которых полностью отсутствуют гистоны, более поздние исследования показали, что их ДНК по-прежнему кодирует гистоновые гены. В отличие от ядерных гистонов, бактериях обнаружены богатые лизином линкерные гистоны (H1), также известные как нуклеопротеины HC1 / HC2.
Архейные гистоны могут быть очень похожи на эволюционных предшественников гистонов эукариот. Гистоновые белкиявляются одними из безопасных белков у эукариот, который укрепляет их роль в биологии ядра. Напротив, зрелые сперматозоиды в основном используют протамины для упаковки своей геномной ДНК, скорее всего потому, что это позволяет достичь еще более высокого коэффициента упаковки.
В некоторых основных формах есть несколько вариантов. классы. Они гомологию аминокислот имеют последовательности и структурное сходство имеют определенные классом основных гистонов, которые отличаются отосновных гистонов. Эти минорные гистоны обычно исполняются функции метаболизма хроматина. Например, гистон H3-подобный CENPA связан только с центром областью хромосомы. Вариант гистона H2A H2A.Z связан с промоторами активно транскрибируемых генов, а также предотвращает распространение молчащего гетерохроматина. Кроме того, H2A.Z играет роль в хроматине для стабильности генома. Другой вариант H2A, H2A.X, фосфорилируется по S139 в областях вокруг двухцепочечныхразрывов и маркирует область, подвергающуюся репарации ДНК. Гистон H3.3 связан с телом активно транскрибируемых генов.
Нуклеосомные (ядерные) гистоны могли развиться из рибосомных белков (RPS6 / RPS15 ), которые имеют они много общего, являясь как короткими, так и одним белками.
Линкерные гистоны имеют гомологи у бактерий.
 Основные структуры хроматина
Основные структуры хроматина Гистоны включают как катушки, вокруг которых наматывается ДНК. Это обеспечивает уплотнение, необходимое для размещения больших геномов эукариот внутри клеток: уплотненная молекула в 40 000 раз короче, чем неупакованная молекула.
 Гистоновые хвосты и функция в образовании хроматина
Гистоновые хвосты и функция в образовании хроматина Гистоны претерпевают посттрансляционные модификации, которые изменяют их взаимодействие с ДНК и ядерными белками. Гистоны H3 и H4 имеютдлинные хвосты, выступающие из нуклеосомы, которые могут быть ковалентно модифицированными в нескольких местах. Модификации хвоста включают метилирование, ацетилирование, фосфорилирование, убиквитинирование, сумоилирование, цитруллинирование и АДФ-рибозилирование. Ядро гистонов H2A и H2B также может быть модифицировано. Считается, что комбинации модификаций составляет код, так называемый «гистоновый код ». Модификации гистонов участвуют в различных биологических процессах, таких как регуляция гена, репарация ДНК, конденсация хромосом (митоз ) и сперматогенез (мейоз ).
Общая номенклатура модификаций гистонов:
Итак H3K4me1 обозначает монометилирование 4- го остатка (лизина) от начала (то есть N-конца ) белка H3.
| Тип. модификации | Гистон | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| H3K4 | H3K9 | H3K14 | H3K27 | H3K79 | H3K36 | H4 K20 | H2BK5 | H2BK20 | |
| моно- метилирование | активация | активация | активация | активация | активация | активация | |||
| диметилирование | репрессия | репрессия | активация | ||||||
| три-метилирование | активация | репрессия | репрессия | активация,. репрессия | активация | репрессия | |||
| ацетилирование | активация | активация | активация | активация | активация | ||||
.
 Схематическое изображение модификаций гистонов. Основано на Родригес-Паредес и Эстеллер, Nature, 2011
Схематическое изображение модификаций гистонов. Основано на Родригес-Паредес и Эстеллер, Nature, 2011 Был описан огромный список модификаций гистонов, но функциональное понимание сообщества из них все еще отсутствует. В совокупности считается, что модификации гистонов могут лежать на основе гистонового кода, в результате чего комбинации модификаций гистонов имеют значения значения. Однако некоторые особенности отдельных заметных модификаций гистонов, которые биохимическиподдаются детальному изучению.
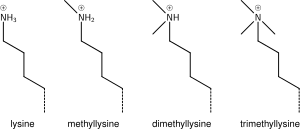
Добавление одной, двух или многих метильных групп к лизину мало влияет на химический состав гистона; Метилирование оставляет лизина нетронутым и оцени минимальное количество элементов, стерические исследования в основном не рассматриваются. Однако белки, содержащие Tudor, хромо или PHD домены, среди прочего, могут распознавать метилирование лизина с исключительной чувствительностью идифференцировать моно, ди и триметиллизин до такой степени, что для лизинов (например, H4K20) моно, ди и три -метилирование, по -видимому, имеет разные значения. Из-за этого метилирование лизина имеет тенденцию быть очень информативной меткой и доминирует над известными функциями модификации гистонов.
Недавно было показано, что добавление группы серотонина в положение 5 глутамина H3 происходит в серотонинергических клетках, таких как нейроны.Это часть дифференцировки серотонинергических клеток. Эта посттрансляционная модификация происходит вместе с модификацией H3K4me3. Серотонилирование усиливает связывание общего фактора транскрипции TFIID с TATA-боксом.
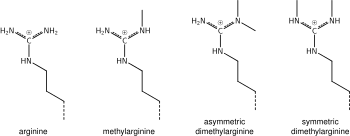
. То, что было сказано выше о химии метилирования лизина, также применимо к метилированию аргинина, а некоторые белковые домены - например, тюдоровские домены - могут быть специфичными для метиларгинина вместо метиллизина. Известно,что аргинин моно- или диметилирован, и метилирование может быть симметричным или асимметричным, своим с разными значениями.
Ферменты, называемые пептидиларгининдезиминазами (PAD), гидролизуют иминную группу аргининов и присоединяют кетогруппу, так что положительный заряд на амино кислотный остаток. Этот процесс активирует экспрессии генов, используемые гистоны менее прочно связанными ДНК и, таким образом, используют хроматин более доступным.PAD также могут иметь противоположный эффект, удаляя или ингибируя моно-метилирование остатков аргинина на гистонах и тем самым препятствиемя положительному эффекту метилирования аргинина на транскрипционную активность.

Добавление ацетильной группы сильное химическое воздействие на лизин, поскольку он нейтрализует положительный заряд. Это снижает электростатическое притяжение между гистоном и отрицательно заряженным острым ДНК, ослабляя хроматина;высокоацетилированные гистоны образуют более доступный хроматин, как правило, связаны с активным транскрипцией. Ацетилирование лизина, по-видимому, имеет меньшее значение, чем метилирование, поскольку гистоновые ацетилтрансферазы имеют тенденцию воздействовать более чем на один лизин; по-предположительно, это должна быть необходимость изменения множества лизинов, чтобы иметь значительный эффект на хроматина. Модификация включает H3K27ac.

Добавление отрицательно заряженной фосфатной группы может привести к серьезным изменениям в структуре белка, что приводит к хорошо изученной роли фосфорилирования в контроле функции белка. Неясно, какие структурные последствия имеет фосфорилирование гистонов, но фосфорилирование гистонов имеет четкие функции как пост-трансляционная модификация, и были охарактеризованы связывающие домены, такие как BRCT.
Наиболее хорошо изученные модификациигистонов участвуют в контроле транскрипции.
Две модификации гистонов особенно связаны с активной транскрипцией:
Три модификации гистонов особенно связаны с репрессированными генами:
Анализ модификаций гистонов в эмбриональных стволовых клетках (и других стволовых клетках) выявил множество промоторов генов, несущих оба H3K4Me3 и H3K27Me3, другими словами, эти промоторы отображают как активирующие, так и репрессирующие метки одновременно. Эта специфическая комбинация модификаций маркирует гены, котор ые готовы к транскрипции; они не требуются в m клеток, но быстро требуются после дифференциации в некоторые клоны. Как только клетка начинаетдифференцироваться, эти двухвалентные промоторы переходят в активное или репрессивное состояние в зависимости от выбранной линии.
Маркировка участков ДНК повреждение является важной функцией модификаций гистонов. Он также защищает ДНК от разрушения ультрафиолетовым излучением солнца.
H3K36me3 обладает способностью рекрутировать комплекс MSH2-MSH6 (hMutSα) пути репарации ошибочного спаривания ДНК. Соответственно, области человеческого генома с высокими уровнями H3K36me3 накапливают меньше соматических мутаций из-за репарации ошибочного спаривания активности.
Epigenet Модификации гистоновых хвостов в определенных областях мозга имеют центральное значение при зависимости. Как только происходят определенные эпигенетические изменения, они, по всей видимости, представляют собой долговечные «молекулярные шрамы», которые могут объяснить стойкость зависимости.
Курильщики сигарет (около 15% населения США) обычно зависимы от никотина.. После 7 дней лечения мышей никотином ацетилирование гистона H3 и гистона H4 увеличивалось на промоторе FosB в ядре . eus accumbens of the brain, causing 61% increase in FosB expression. This would also increase expression of the splice variant Delta FosB. In the nucleus accumbens of the brain, Delta FosB functions as a "sustained molecular switch" and "master control protein" in the development of an addiction.
About 7% of the US population is addicted to alcohol. In rats exposed to alcohol for up to 5 days, there was an increase in histone 3 lysine 9ацетилирование в промоторе пронцицептина в комплексе миндалины мозга. Это ацетилирование является активирующей меткой для пронцицептина. Система опиоидных рецепторов ноцицептина / ноцицептина участвует в усиливающих или кондиционирующих эффектах алкоголя.
Метамфетаминовая зависимость встречается примерно у 0,2% населения США. Хроническое употребление метамфетамина вызывает метилирование лизина в положении 4 гистона 3, расположенного на промоторах c-fos и C-C гены хемокинового рецептора 2 (ccr2), активирующие эти гены в прилежащем ядре (NAc). c-fos, как известно, играет важную роль в зависимости. Ген ccr2 также важен при зависимости, поскольку мутационная инактивация этого гена снижает зависимость.
Первым шагом дупликации структуры хроматина является синтез гистоновых белков: H1, H2A, H2B, H3, H4. Эти белки синтезируются во время фазы S клеточного цикла. Есть разные мехи механизмы, которые способствуют увеличению синтеза гистонов.
Дрожжи несут одну или две копии каждого гистонового гена, которые не сгруппированы, а скорее разбросаны по хромосомам. Транскрипция гена гистона контролируется множеством регуляторных белков генов, таких как факторы транскрипции, которые связываются с участками промотора гистонов. В почкующихся дрожжах ген-кандидат для активации экспрессии гистонового гена - SBF. SBF - фактор транскрипции, который активируется в конце Фаза G1, когда она отделяется от своего репрессора Whi5. Это происходит, когда Whi5 фосфорилируется Cdc8, который является G1 / S Cdk. Подавление экспрессии гистонового гена вне S-фаз зависит от белков Hir, которые образуют неактивную структуру хроматина в локусе гистоновых генов, вызывая блокировку активаторов транскрипции. в скорости синтеза гистонов происходит за счет увеличения процессинга пре-мРНК до ее мат форма мочевины, а также уменьшение деградации мРНК; это приводит к увеличению активной мРНК для трансляции гистоновых белков. Было обнаружено, что механизм активации мРНК заключается в удалении сегмента на 3'-конце цепи мРНК и зависит от ассоциации с белком, связывающим стержень-петлю (SLBP ). SLBP также стабилизирует мРНК гистонов во время S фазы, блокируя деградацию нуклеазой 3'hExo. Уровни SLBP контролируются белками клеточного цикла, заставляя SLBP накапливаться, когда клетки входят в S-фазу, и деградировать, когда клетки покидают S-фазу. SLBP маркируются для деградации путем фосфорилирования по двум остаткам треонина циклин-зависимыми киназами, возможно, циклином A / cdk2, в конце S-фазы. У многоклеточных животных также есть несколько копий гистоновых генов, сгруппированных на хромосомах, которые локализованы в структурах, называемых тельцами Кахаля, что определяется анализом захвата конформации хромосомы по всему геному (4C-Seq).
Ядерный белок атаксии-телеангиэктазии (NPAT), также известный как ядерный белок-коактиватор транскрипции гистонов, представляет собой фактор транскрипции, который активирует транскрипцию гена гистона на хромосомах 1 и 6 клеток человека. NPAT также является субстрат циклина E-Cdk2, который необходим для перехода между фазой G1 и фазой S. NPAT активирует экспрессию гистонового гена только после того, как он былфосфорилирован G1 / S-Cdk циклином E-Cdk2 в ранней S-фазе. Это показывает регуляторную связь между контролем клеточного цикла и синтезом гистонов.
| Найдите гистон вВикисловаре, бесплатном формате. |