
Войти
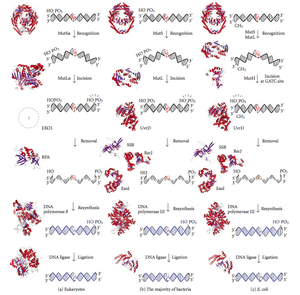 Схема путей восстановления несоответствия ДНК. Первый столбец показывает восстановление несоответствия у эукариот, а второй - восстановление у большинства бактерий. В третьем столбце показано исправление несоответствия, а именно в E. coli.
Схема путей восстановления несоответствия ДНК. Первый столбец показывает восстановление несоответствия у эукариот, а второй - восстановление у большинства бактерий. В третьем столбце показано исправление несоответствия, а именно в E. coli.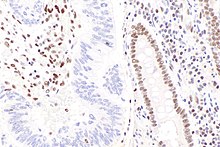 Микрофотография, показывающая потерю окрашивания на MLH1 в аденокарциноме прямой кишки в соответствии с репарацией несоответствия ДНК (слева от изображения) и доброкачественной слизистой оболочки толстой кишки (справа от изображения).
Микрофотография, показывающая потерю окрашивания на MLH1 в аденокарциноме прямой кишки в соответствии с репарацией несоответствия ДНК (слева от изображения) и доброкачественной слизистой оболочки толстой кишки (справа от изображения). Восстановление несоответствия ДНК (MMR ) - это система для распознавания и исправления ошибочной вставки, удаления и неправильного включения оснований, которые могут возникнуть во время репликации ДНК и рекомбинация, а также восстановление некоторых форм повреждения ДНК.
Ремонт неправильного спаривания является нитьспецифичным. Во время синтеза ДНК вновь синтезированная (дочерняя) цепь обычно будет содержать ошибки. Чтобы начать восстановление, механизм восстановления несоответствия отличает вновь синтезированную цепь от шаблона (родительского). У грамотрицательных бактерий временное гемиметилирование различает цепи (родительский элемент метилирован, а дочерний - нет). Однако у других прокариот и эукариот точный механизм не ясен. Предполагается, что у эукариот вновь синтезированная ДНК с отстающей цепью временно содержит зарубки (до того, как она запечатывается ДНК-лигазой) и обеспечивает сигнал, который направляет системы проверки несоответствия на соответствующую цепь. Это означает, что эти зазубрины должны присутствовать в ведущей нити, и недавно были обнаружены доказательства этого. Недавняя работа показала, что зарубки являются сайтами для RFC-зависимой загрузки репликационного скользящего зажима PCNA специфическим для ориентации образом, так что одна сторона белка в форме пончика соприкасается с 3'-OH концом на зазубрине. Затем загруженная PCNA направляет действие эндонуклеазы MutLalpha на дочернюю цепь в присутствии несоответствия и MutSalpha или MutSbeta.
Любое мутационное событие, которое нарушает сверхспиральную структуру ДНК, несет в себе возможность поставить под угрозу генетическую стабильность клетки. Тот факт, что системы обнаружения и восстановления повреждений столь же сложны, как и сам механизм репликации, подчеркивает важность, которую эволюция придавала точности ДНК.
Примеры несовпадающих оснований включают спаривание G / T или A / C (см. репарация ДНК ). Несоответствия обычно возникают из-за таутомеризации оснований во время репликации ДНК. Повреждение восстанавливается путем распознавания деформации, вызванной несоответствием, определения матрицы и нити, не являющейся шаблоном, и удаления неправильно встроенного основания и замены его правильным нуклеотидом. Процесс удаления включает в себя не только сам несовпадающий нуклеотид. Может быть удалено несколько или до тысяч пар оснований вновь синтезированной цепи ДНК.
| Белок репарации несоответствия ДНК, С-концевой домен | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 hpms2-atpgs hpms2-atpgs | |||||||||
| Идентификаторы | |||||||||
| Символ | DNA_mis_repair | ||||||||
| Pfam | PF01119 | ||||||||
| Pfam клан | CL0329 | ||||||||
| InterPro | IPR013507 215>PROSITE | PDOC00057 | |||||||
| SCOPe | 1bkn / SUPFAM | ||||||||
| |||||||||
Исправление несоответствия - это высококонсервативный процесс от прокариот до эукариот. Первые доказательства исправления несоответствия были получены от S. pneumoniae (гены hexA и hexB ). Последующие работы по Э. coli идентифицировал ряд генов, которые при мутационной инактивации вызывают гипермутационные штаммы. Поэтому генные продукты называются «Mut» белками и являются основными активными компонентами системы репарации ошибочного спаривания. Три из этих белков важны для обнаружения несоответствия и направления к нему механизма репарации: MutS, MutH и MutL (MutS является гомологом HexA и MutL HexB).
MutS образует димер (MutS 2), который распознает несовпадающее основание на дочерней цепи и связывает мутировавшую ДНК. MutH связывается в гемиметилированных сайтах дочерней ДНК, но его действие является латентным, активируясь только при контакте с димером MutL (MutL 2), который связывает комплекс MutS-ДНК и действует как посредник между MutS 2 и MutH, активируя последний. ДНК зацикливается для поиска ближайшего сайта метилирования d (GATC) к несоответствию, который может находиться на расстоянии до 1 т.п.н. После активации комплексом MutS-ДНК MutH разрывает дочернюю цепь рядом с гемиметилированным сайтом. MutL рекрутирует UvrD геликазу (ДНК-геликаза II) для разделения двух цепей с определенной полярностью от 3 'до 5'. Затем весь комплекс MutSHL скользит по ДНК в направлении несовпадения, высвобождая цепь, которую нужно вырезать по ходу движения. Экзонуклеаза следует за комплексом и переваривает хвост ss-ДНК. Привлечение экзонуклеазы зависит от того, с какой стороны несовпадающего MutH надрезает цепь - 5 'или 3'. Если ник, сделанный MutH, находится на 5'-конце несоответствия, используется либо RecJ, либо ExoVII (обе экзонуклеазы от 5 'до 3'). Если, однако, зарубка находится на 3 'конце несовпадения, используется (фермент от 3' до 5 ').
Весь процесс заканчивается за сайтом несоответствия, т.е. как сам сайт, так и окружающие его нуклеотиды полностью вырезаются. Одноцепочечный разрыв, созданный экзонуклеазой, затем может быть восстановлен ДНК-полимеразой III (с помощью одноцепочечного связывающего белка), который использует другую цепь в качестве матрицы и, наконец, запечатан ДНК-лигазой. Затем ДНК-метилаза быстро метилирует дочернюю цепь.
При связывании димер MutS 2 изгибает спираль ДНК и экранирует приблизительно 20 пар оснований. Он обладает слабой АТФазной активностью, и связывание АТФ приводит к образованию третичных структур на поверхности молекулы. Кристаллическая структура MutS показывает, что она исключительно асимметрична, и, хотя его активная конформация представляет собой димер, только одна из двух половин взаимодействует с сайтом несовпадения.
У эукариот омологи MutShобразуют два основных гетеродимера: Msdiv class="ht" / Msh6 (MutSα) и Msdiv class="ht" / Msh3 (MutSβ). Путь MutSα участвует в основном в замене оснований и репарации ошибочного спаривания с малой петлей. Путь MutSβ также участвует в репарации малой петли в дополнение к репарации большой петли (~ 10 нуклеотидных петель). Однако MutSβ не восстанавливает замены оснований.
MutL также обладает слабой АТФазной активностью (он использует АТФ для движения). Он образует комплекс с MutS и MutH, увеличивая след MutS в ДНК.
Однако процессивность (расстояние, на которое фермент может пройти по ДНК до диссоциации) UvrD составляет всего ~ 40-50 п.н. Поскольку расстояние между ником, созданным MutH, и несоответствием может составлять в среднем ~ 600 п.н., если другой UvrD не загружен, размотанная секция может повторно отжигаться до своей дополнительной цепи, заставляя процесс начинаться заново. Однако при помощи MutL скорость загрузки UvrD значительно увеличивается. Хотя процессивность (и использование АТФ) отдельных молекул UvrD остается неизменной, общее воздействие на ДНК значительно усиливается; ДНК не имеет шанса на повторный отжиг, поскольку каждый UvrD раскручивает 40-50 п.н. ДНК, диссоциирует, а затем немедленно заменяется другим UvrD, повторяя процесс. Это подвергает большие участки ДНК перевариванию экзонуклеазой, что позволяет быстро вырезать (и позднее заменить) неправильную ДНК.
У эукариот есть пять MutLhомологов, обозначенных как MLH1, MLH2, MLH3, PMS1 и PMS2. Они образуют гетеродимеры, имитирующие MutL в E. coli. Человеческие гомологи прокариотического MutL образуют три комплекса, обозначаемых как MutLα, MutLβ и MutLγ. Комплекс MutLα состоит из субъединиц MLH1 и PMS2, гетеродимер MutLβ состоит из MLH1 и PMS1, тогда как MutLγ состоит из MLH1 и MLH3. MutLα действует как эндонуклеаза, которая вызывает разрывы цепи в дочерней цепи при активации ошибочным спариванием и другими необходимыми белками, MutSα и PCNA. Эти прерывания цепи служат точками входа для экзонуклеазной активности, которая удаляет несовпадающую ДНК. Роли, которые играют MutLβ и MutLγ в репарации несоответствий, менее понятны.
MutH - это очень слабая эндонуклеаза, которая активируется после связывания с MutL (который сам связан с MutS). Он разрывает неметилированную ДНК и неметилированную цепь гемиметилированной ДНК, но не разрывает полностью метилированную ДНК. Эксперименты показали, что восстановление несоответствия происходит случайно, если ни одна из цепей не метилирована. Такое поведение привело к предположению, что MutH определяет, какая нить содержит несоответствие. MutH не имеет эукариотического гомолога. Его эндонуклеазная функция выполняется гомологами MutL, которые обладают некоторой специализированной 5'-3'-экзонуклеазной активностью. Смещение цепи для удаления несовпадений из вновь синтезированной дочерней цепи у эукариот может быть обеспечено свободными 3'-концами фрагментов Окадзаки в новой цепи, созданной во время репликации.
PCNA и β-скользящий зажим связаны с MutSα / β и MutS соответственно. Хотя первоначальные сообщения предполагали, что комплекс PCNA-MutSα может усиливать распознавание ошибочного спаривания, недавно было продемонстрировано, что нет явного изменения аффинности MutSα к несовпадению в присутствии или в отсутствие PCNA. Кроме того, мутанты MutSα, которые не могут взаимодействовать с PCNA in vitro, проявляют способность осуществлять распознавание несоответствия и удаление несоответствия до уровней, близких к дикому типу. Такие мутанты являются дефектными в реакции репарации, направляемой разрывом 5'-цепи, что впервые указывает на функцию MutSα на стадии реакции после эксцизии.
Мутации в человеческих гомологах белков Mut влияют на стабильность генома, что может привести к микросателлитной нестабильности (MSI), причастный к некоторым раковым заболеваниям человека. В частности, наследственный неполипозный колоректальный рак (HNPCC или синдром Линча) объясняется повреждением вариантов зародышевой линии в генах, кодирующих гомологи MutS и MutL MSH2 и MLH1 соответственно, которые классифицируются как гены-супрессоры опухолей. Один подтип HNPCC, синдром Мюира-Торре (MTS), связан с опухолями кожи. Если обе унаследованные копии (аллели) гена MMR несут повреждающие генетические варианты, это приводит к очень редкому и тяжелому состоянию: синдром репарации несоответствия (или конституциональный дефицит репарации несоответствия, CMMR-D), проявляющийся как множественные опухоли в раннем возрасте, часто опухоли толстой кишки и головного мозга.
Спорадические виды рака с дефицитом репарации ДНК очень редко имеют мутацию в гене репарации ДНК, но вместо этого они, как правило, имеют эпигенетические изменения, такие как метилирование промотора, которые ингибируют экспрессию гена репарации ДНК. Около 13% случаев колоректального рака не имеют возможности репарации несоответствия ДНК, обычно из-за потери MLH1 (9,8%) или иногда MSH2, MSH6 или PMS2 (все ≤1,5%). Для большинства спорадических случаев колоректального рака с дефицитом MLH1 этот дефицит был вызван метилированием промотора MLH1. Другие типы рака имеют более высокие частоты потери MLH1 (см. Таблицу ниже), что опять же в значительной степени является результатом метилирования промотора гена MLH1. Другой эпигенетический механизм, лежащий в основе дефицита MMR, может включать сверхэкспрессию микроРНК, например, уровни miR-155 обратно коррелируют с экспрессией MLH1 или MSH2 при колоректальном раке.
| Тип рака | Частота дефицита при раке | Частота дефицита в соседнем полевом дефекте |
|---|---|---|
| Желудок | 32% | 24% - 28% |
| Желудок (опухоли фовеолярного типа) | 74% | 71% |
| Желудок в долине Кашмира с высокой заболеваемостью | 73% | 20% |
| пищевода | 73% | 27% |
| плоскоклеточный рак головы и шеи (HNSCC) | 31% -33% | 20% -25% |
| Немелкоклеточный рак легкого (НМРЛ) | 69% | 72% |
| Колоректальный | 10% |
A полевые дефекты (полевое канцерирование) - это область эпителия, которая была обусловлена эпигенетическими или генетическими изменениями, предрасполагающими к развитию рака. Как указывает Рубин, «... есть свидетельства того, что более 80% соматических мутаций, обнаруживаемых в опухолях толстой кишки человека мутаторного фенотипа, происходят до начала терминальной клональной экспансии». Аналогичным образом Vogelstein et al. указывают на то, что более половины соматических мутаций, выявленных в опухолях, произошли в предопухолевой фазе (в области дефекта поля), во время роста явно нормальных клеток.
Дефицит MLH1 был обычным явлением в полевых дефектах (гистологически нормальных тканях), окружающих опухоли; см. таблицу выше. Эпигенетически замалченный или мутированный MLH1, вероятно, не предоставит селективного преимущества стволовым клеткам, однако это вызовет повышенную частоту мутаций, и один или несколько мутировавших генов могут обеспечить клетке селективное преимущество. Дефицитный ген MLH1 может затем переноситься в качестве избирательно почти нейтрального гена-пассажира (автостопщика), когда мутировавшая стволовая клетка генерирует расширенный клон. Продолжающееся присутствие клона с эпигенетически репрессированным MLH1 будет продолжать генерировать дальнейшие мутации, некоторые из которых могут привести к опухоли.
У людей семь белков восстановления несоответствия ДНК (MMR) (MLH1, MLH3, MSH2, MSH3, MSH6, PMS1 и PMS2 ) работают скоординированно в последовательных этапах, чтобы инициировать восстановление несоответствий ДНК. Кроме того, существуют Exo1 -зависимые и Exo1-независимые подпатки MMR.
Другие генные продукты, участвующие в репарации несоответствия (после инициации генами MMR) у людей, включают ДНК-полимеразу дельта, PCNA, RPA, HMGB1, RFC и ДНК-лигаза I, плюс факторы, модифицирующие гистон и хроматин.
При определенных обстоятельствах путь MMR может задействовать подверженную ошибкам ДНК-полимеразу eta (POLH ). Это происходит в B-лимфоцитах во время соматической гипермутации, где POLH используется для введения генетической изменчивости в гены антител. Однако этот подверженный ошибкам путь MMR может запускаться в других типах клеток человека при воздействии генотоксинов, и, действительно, он широко активен при различных раковых заболеваниях человека, вызывая мутации, несущие сигнатуру активности POLH.
Распознавание и устранение несовпадений и инделек важно для клеток, поскольку невыполнение этого приводит к микросателлитной нестабильности (MSI) и повышенной скорости спонтанных мутаций (мутатор фенотип). По сравнению с другими типами рака, рак с дефицитом MMR (MSI) имеет очень высокую частоту мутаций, близких к меланоме и раку легких, типам рака, вызванным сильным воздействием УФ-излучения и мутагенных химических веществ.
В дополнение к очень высокому бремени мутаций, дефицит MMR приводит к необычному распределению соматических мутаций по геному человека: это говорит о том, что MMR предпочтительно защищает богатые генами, рано реплицирующиеся эухроматические области. Напротив, бедные генами, поздно реплицирующиеся гетерохроматические области генома демонстрируют высокую частоту мутаций во многих опухолях человека.
модификация гистона H3K36me3, эпигенетическая метка активного хроматина, обладает способностью рекрутировать комплекс MSH2-MSH6 (hMutSα). Соответственно, участки генома человека с высоким уровнем H3K36me3 накапливают меньше мутаций из-за активности MMR.
Отсутствие MMR часто происходит в координации с потерей MMR. другие гены репарации ДНК. Например, гены MMR MLH1 и MLH3, а также 11 других генов репарации ДНК (таких как MGMT и многие гены пути NER ) были значительно подавлены как при астроцитомах более низкого, так и при более высоком уровне, в отличие от нормальной ткани мозга. Более того, экспрессия MLH1 и MGMT была тесно коррелирована в 135 образцах рака желудка, а потеря MLH1 и MGMT, по-видимому, синхронно ускорялась во время прогрессирования опухоли.
Недостаточная экспрессия нескольких генов репарации ДНК часто обнаруживается при раке и может вносить вклад в тысячи мутаций, обычно обнаруживаемых при раке (см. Частота мутаций при раке ).