
Войти
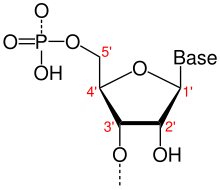 A фуранозы (сахарное кольцо) с атомами углерода, помеченными с использованием стандартных обозначений. 5 'находится выше по потоку; 3 'находится ниже по потоку. ДНК и РНК синтезируются в направлении от 5 'к 3'.
A фуранозы (сахарное кольцо) с атомами углерода, помеченными с использованием стандартных обозначений. 5 'находится выше по потоку; 3 'находится ниже по потоку. ДНК и РНК синтезируются в направлении от 5 'к 3'. Направленность, в молекулярной биологии и биохимии, представляет собой химическую ориентацию от конца к концу одиночной цепи нуклеиновой кислоты. В одной цепи ДНК или РНК химическое соглашение об именах атомов углерода в нуклеотиде сахарном кольце означает, что будет быть 5 '-концом (обычно произносится как «пять простых концов»), который часто содержит фосфатную группу, присоединенную к 5'-углеродному циклу рибозного кольца. и 3 '-конец (обычно произносится как «три простых конца»), который обычно немодифицирован по отношению к заместителю рибозы -ОН. В двойной спирали ДНК, нити проходят в противоположных направлениях, чтобы между ними было возможно спаривание оснований, что важно для репликации или транскрипции кодированной информации.
Нуклеиновые кислоты могут быть синтезированы in vivo только в направлении от 5'-к-3 ', поскольку полимеразы, которые собирают различные типы новых цепей, обычно зависят от энергия, полученная при разрыве нуклеозидтрифосфатных связей для присоединения новых нуклеозидных монофосфатов к 3'- гидроксильной (-OH) группе через фосфодиэфирную связь. Относительные положения структур вдоль цепи нуклеиновой кислоты, включая гены и различные белковые сайты связывания, обычно отмечаются как расположенные выше (к 5'-концу) или ниже по течению ( ближе к 3′-концу). (См. Также восходящий и нисходящий.)
Направленность связана с смыслом, но не зависит от него. Транскрипция одноцепочечной РНК из двухцепочечной матрицы ДНК требует выбора одной цепи матрицы ДНК в качестве цепи матрицы, которая напрямую взаимодействует с формирующейся РНК благодаря комплементарной последовательности. Другая цепь не копируется напрямую, но обязательно ее последовательность будет аналогична последовательности РНК. Сайты инициации транскрипции обычно встречаются на обеих цепях ДНК организма и определяют местоположение, направление и обстоятельства, при которых будет происходить транскрипция. Если транскрипт кодирует один или (редко) несколько белков, трансляция каждого белка с помощью рибосомы будет происходить в направлении от 5 'до 3' и будет вытягивать белок от его N конец к его концу C. Например, в типичном гене стартовый кодон (5'-ATG-3 ') представляет собой последовательность ДНК в смысловой цепи. Транскрипция начинается с вышестоящего сайта (относительно смысловой цепи), и по мере прохождения через область она копирует 3'-TAC-5 'из цепочки-матрицы с образованием 5'-AUG-3' в мессенджере . РНК (мРНК). МРНК сканируется рибосомой с 5'-конца, где стартовый кодон направляет включение метионина (бактерии, митохондрии и пластиды используют N-формилметионин вместо) на N-конце белка. По соглашению, одиночные цепи последовательностей ДНК и РНК записываются в направлении от 5'-к-3 ', за исключением случаев, когда это необходимо для иллюстрации модели спаривания оснований.
 В Показанный сегмент ДНК, направления от 5 'к 3' идут вниз по левой цепи и вверх по правой цепи
В Показанный сегмент ДНК, направления от 5 'к 3' идут вниз по левой цепи и вверх по правой цепи 5'-конец (произносится как «пять основных концов») обозначает конец цепи ДНК или РНК, имеющей пятую цепь. углерод в сахарном кольце дезоксирибозы или рибозы на его конце. фосфатная группа, присоединенная к 5'-концу, допускает лигирование двух нуклеотидов, т.е. ковалентное связывание 5'-фосфата с 3'- гидроксильная группа другого нуклеотида с образованием фосфодиэфирной связи. Удаление 5'-фосфата предотвращает лигирование. Чтобы предотвратить нежелательное лигирование нуклеиновых кислот (например, самолигирование плазмидного вектора в клонирование ДНК ), молекулярные биологи обычно удаляют 5'-фосфат с помощью фосфатаза.
5'-конец формирующейся матричной РНК представляет собой сайт, на котором происходит посттранскрипционное кэппирование, процесс, который имеет жизненно важное значение для получения зрелой матричной РНК. Кеппинг увеличивает стабильность матричной РНК, пока она подвергается трансляции, обеспечивая устойчивость к разрушающим эффектам экзонуклеаз. Он состоит из метилированного нуклеотида (метилгуанозин ), присоединенного к матричной РНК с помощью редкой 5'- к 5'-трифосфатной связи.
5'-фланкирующая область гена часто обозначает область ДНК, которая не транскрибируется в РНК. 5'-фланкирующая область содержит промотор гена , а также может содержать энхансеры или другие сайты связывания с белками.
5'-нетранслируемая область (5'-UTR) представляет собой область гена, которая транскрибируется в мРНК и расположена на 5'-конце мРНК. Эта область мРНК может быть, а может и не быть транслирована, но обычно участвует в регуляции трансляции. 5'-нетранслируемая область представляет собой часть ДНК, начинающуюся от кэп-сайта и простирающуюся до основания непосредственно перед кодоном инициации трансляции AUG основной кодирующей последовательности. Эта область может иметь последовательности, такие как сайт связывания рибосомы и последовательность Козака, которые определяют эффективность трансляции мРНК или которые могут влиять на стабильность мРНК.
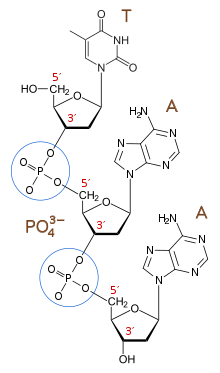 Фосфодиэфирные связи (обведены) между нуклеотидами
Фосфодиэфирные связи (обведены) между нуклеотидами 3'-конец (три простых конца) цепи назван так из-за того, что он оканчивается на гидроксильная группа третьего углерода в сахарном кольце и известна как хвостовой конец. 3'-гидроксил необходим в синтезе новых молекул нуклеиновой кислоты, так как он лигирован (присоединен) к 5'-фосфату отдельного нуклеотида, что позволяет образовывать цепи связанных нуклеотидов.
Молекулярные биологи могут использовать нуклеотиды, в которых отсутствует 3'-гидроксил (дидезоксирибонуклеотиды), для прерывания репликации ДНК. Этот метод известен как метод завершения дидезокси-цепи или метод Сэнгера, и используется для определения порядка нуклеотидов в ДНК..
3'-конец зарождающейся матричной РНК является сайт посттранскрипционного полиаденилирования, который присоединяет цепь из 50-250 аденозиновых остатков для образования зрелой матричной РНК. Эта цепочка помогает определить, как долго информационная РНК существует в клетке, влияя на то, сколько из нее вырабатывается белка.
3'-фланкирующая область представляет собой область ДНК, которая не копируется в зрелую мРНК, но которая присутствует рядом с 3'-концом гена. Первоначально считалось, что 3'-фланкирующая ДНК вообще не транскрибируется, но было обнаружено, что она транскрибируется в РНК и быстро удаляется во время обработки первичного транскрипта с образованием зрелой мРНК. 3'-фланкирующая область часто содержит последовательности, которые влияют на формирование 3'-конца сообщения. Он также может содержать энхансеры или другие сайты, с которыми могут связываться белки.
3'-нетранслируемая область (3'-UTR) - это область ДНК, которая транскрибируется в мРНК и становится 3'-концом сообщения, но не содержат последовательность, кодирующую белок. Все, что находится между стоп-кодоном и polyA-хвостом, считается 3'-нетранслированным. 3'-нетранслируемая область может влиять на эффективность трансляции мРНК или стабильность мРНК. Он также имеет последовательности, которые требуются для добавления поли (A) хвоста к сообщению, включая гексануклеотид AAUAAA.