
Войти
 Основные единицы структуры хроматина
Основные единицы структуры хроматина Нуклеосом является основным структурным звеном ДНК упаковки в эукариот. Структура нуклеосомы состоит из сегмента ДНК, намотанного вокруг восьми гистоновых белков, и напоминает нить, намотанную на катушку. Нуклеосома - основная субъединица хроматина. Каждая нуклеосома состоит из немногим менее двух витков ДНК, обернутых вокруг набора из восьми белков, называемых гистонами, которые известны как октамер гистонов. Каждый октамер гистонов состоит из двух копий каждого из гистоновых белков H2A, H2B, H3 и H4.
ДНК должна быть уплотнена в нуклеосомы, чтобы соответствовать ядру клетки. Помимо обертывания нуклеосом, хроматин эукариот дополнительно уплотняется, складываясь в ряд более сложных структур, в конечном итоге формируя хромосому. Каждая клетка человека содержит около 30 миллионов нуклеосом.
Считается, что нуклеосомы несут эпигенетически унаследованную информацию в форме ковалентных модификаций своих основных гистонов. Положения нуклеосом в геноме не случайны, и важно знать, где находится каждая нуклеосома, потому что это определяет доступность ДНК для регуляторных белков.
Нуклеосомы были впервые обнаружены как частицы в электронном микроскопе Доном и Адой Олинс в 1974 году, а их существование и структура (в виде октамеров гистонов, окруженных приблизительно 200 парами оснований ДНК) были предложены Роджером Корнбергом. Роль нуклеосомы как общего репрессора генов была продемонстрирована Lorch et al. in vitro и Han и Grunstein in vivo в 1987 и 1988 годах соответственно.
Частица ядра нуклеосомы состоит из приблизительно 146 пар оснований (п.н.) ДНК, обернутых 1,67 левых сверхспиральных витков вокруг октамера гистонов, состоящих из 2 копий каждого из гистонов ядра H2A, H2B, H3 и H4. Частицы ядра связаны отрезками линкерной ДНК, длина которых может составлять до 80 п.н. Технически нуклеосома определяется как коровая частица плюс одна из этих линкерных областей; однако это слово часто является синонимом основной частицы. Полногеномные карты расположения нуклеосом теперь доступны для многих модельных организмов, включая печень и мозг мышей.
Линкерные гистоны, такие как H1 и его изоформы, участвуют в уплотнении хроматина и сидят в основании нуклеосомы рядом с входом и выходом ДНК, связываясь с линкерной областью ДНК. Неконденсированные нуклеосомы без линкерного гистона напоминают «бусинки на нити ДНК» под электронным микроскопом.
В отличие от большинства эукариотических клеток, зрелые сперматозоиды в основном используют протамины для упаковки своей геномной ДНК, что, скорее всего, обеспечивает еще более высокий коэффициент упаковки. Эквиваленты гистонов и упрощенная структура хроматина также были обнаружены у архей, что позволяет предположить, что эукариоты - не единственные организмы, использующие нуклеосомы.
 Кристаллическая структура ядерной частицы нуклеосомы, состоящая из ядерных гистонов H2A, H2B, H3 и H4 и ДНК. Вид сверху через сверхспиральную ось.
Кристаллическая структура ядерной частицы нуклеосомы, состоящая из ядерных гистонов H2A, H2B, H3 и H4 и ДНК. Вид сверху через сверхспиральную ось. Пионерские структурные исследования в 1980-х годах, проведенные группой Аарона Клуга, предоставили первое свидетельство того, что октамер гистоновых белков оборачивает ДНК вокруг себя примерно на 1,7 витка левой суперспирали. В 1997 году группа Ричмонда решила первую кристаллическую структуру нуклеосомы с близким к атомному разрешению, показав наиболее важные детали частицы. Человеческий альфа - спутник палиндромных ДНК решающих значения для достижения в 1997 году нуклеосом кристаллической структуры были разработаны группой Bunick в Oak Ridge National Laboratory в Теннесси. На сегодняшний день решены структуры более 20 различных ядер нуклеосомных частиц, включая те, которые содержат гистоновые варианты и гистоны разных видов. Структура ядерной частицы нуклеосомы удивительно консервативна, и даже изменение более чем на 100 остатков между гистонами лягушки и дрожжей приводит к картам электронной плотности с общим среднеквадратичным отклонением всего 1,6 Å.
Частица ядра нуклеосомы (показанная на рисунке) состоит из примерно 146 пар оснований ДНК, обернутых 1,67 левых сверхспиральных витков вокруг октамера гистонов, состоящих из 2 копий каждого из гистонов ядра H2A, H2B, H3 и H4. Соседние нуклеосомы соединены участком свободной ДНК, называемым линкерной ДНК (длина которого варьируется от 10 до 80 п.н. в зависимости от вида и типа ткани). Вся структура образует цилиндр диаметром 11 нм и высотой 5,5 нм.
 Апоптотическая лестница ДНК. Переваренный хроматин находится на первой полосе; второй содержит стандарт ДНК для сравнения длин.
Апоптотическая лестница ДНК. Переваренный хроматин находится на первой полосе; второй содержит стандарт ДНК для сравнения длин. 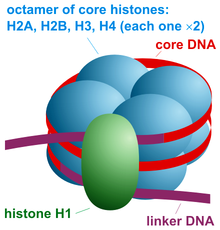 Схема организации нуклеосом.
Схема организации нуклеосом.  Кристаллическая структура ядерной частицы нуклеосомы ( PDB : 1EQZ )
Кристаллическая структура ядерной частицы нуклеосомы ( PDB : 1EQZ ) Частицы ядра нуклеосомы наблюдаются, когда хроматин в интерфазе обрабатывают, чтобы заставить хроматин частично разворачиваться. Полученное изображение, полученное с помощью электронного микроскопа, представляет собой «бусинки на нитке». Нить - это ДНК, а каждая бусинка в нуклеосоме - это центральная частица. Частица ядра нуклеосомы состоит из ДНК и гистоновых белков.
При частичном расщеплении хроматина ДНКазой обнаруживается его нуклеосомная структура. Поскольку участки ДНК ядер нуклеосомных частиц менее доступны для ДНКазы, чем связывающие участки, ДНК расщепляется на фрагменты, длина которых равна кратности расстояния между нуклеосомами (180, 360, 540 пар оснований и т. Д.). Следовательно, во время гель-электрофореза этой ДНК виден очень характерный узор, похожий на лестницу. Такое переваривание может происходить также в естественных условиях во время апоптоза («клеточное самоубийство» или запрограммированная гибель клеток), поскольку его роль обычно заключается в саморазрушении ДНК.
Основные гистоновые белки содержат характерный структурный мотив, называемый «гистоновая складка», который состоит из трех альфа-спиралей (α1-3), разделенных двумя петлями (L1-2). В растворе гистоны образуют гетеродимеры H2A-H2B и гетеротетрамеры H3-H4. Гистоны димеризуются вокруг своих длинных α2 спиралей в антипараллельной ориентации, и в случае H3 и H4 два таких димера образуют 4-спиральный пучок, стабилизированный обширным взаимодействием H3-H3 '. Димер H2A / H2B связывается с тетрамером H3 / H4 за счет взаимодействий между H4 и H2B, которые включают образование гидрофобного кластера. Октамер гистонов образован центральным тетрамером H3 / H4, зажатым между двумя димерами H2A / H2B. Из-за высокого основного заряда всех четырех ядер гистонов октамер гистонов стабилен только в присутствии ДНК или очень высоких концентраций соли.
Нуклеосома содержит более 120 прямых взаимодействий белок-ДНК и несколько сотен опосредованных водой. Прямые взаимодействия белок-ДНК не распределяются равномерно по поверхности октамера, а скорее расположены в дискретных участках. Это происходит из-за образования двух типов сайтов связывания ДНК внутри октамера; сайт α1α1, который использует спираль α1 из двух соседних гистонов, и сайт L1L2, образованный петлями L1 и L2. Солевые связи и водородные связи между основной и гидроксильной группами боковой цепи и амидами основной цепи с фосфатами основной цепи ДНК образуют основную часть взаимодействий с ДНК. Это важно, учитывая, что повсеместное распределение нуклеосом по геномам требует, чтобы он был неспецифическим для последовательности ДНК-связывающим фактором. Хотя нуклеосомы, как правило, предпочитают одни последовательности ДНК другим, они способны связываться практически с любой последовательностью, что, как считается, связано с гибкостью в образовании этих опосредованных водой взаимодействий. Кроме того, неполярные взаимодействия происходят между боковыми цепями белка и дезоксирибозными группами, и боковая цепь аргинина внедряется в малую бороздку ДНК во всех 14 участках, где она обращена к поверхности октамера. Распределение и сила ДНК-связывающих сайтов на поверхности октамера искажает ДНК в ядре нуклеосомы. ДНК неравномерно изогнута, а также содержит дефекты скручивания. Скручивание свободной B-формы ДНК в растворе составляет 10,5 п.н. за оборот. Однако общий поворот нуклеосомной ДНК составляет всего 10,2 п.н. на оборот, варьируя от 9,4 до 10,9 п.н. на оборот.
Удлинения гистоновых хвостов составляют до 30% по массе гистонов, но не видны в кристаллических структурах нуклеосом из-за их высокой внутренней гибкости и, как полагают, в значительной степени неструктурированы. N-концевые хвосты гистонов H3 и H2B проходят через канал, образованный малыми бороздками двух нитей ДНК, выступающими из ДНК каждые 20 п.н. N-концевой хвост гистона Н4, с другой стороны, имеет область высокообогащенного основных аминокислот (16-25), который, в кристаллической структуре, образует взаимодействие с сильнокислой области поверхности димера Н2А-Н2В другой нуклеосомы, потенциально имеющей отношение к структуре нуклеосом более высокого порядка. Это взаимодействие, как полагают, также происходит в физиологических условиях и предполагает, что ацетилирование хвоста H4 искажает структуру хроматина более высокого порядка.
 Текущая модель уплотнения хроматина.
Текущая модель уплотнения хроматина. Организация ДНК, которая достигается нуклеосомой, не может полностью объяснить упаковку ДНК, наблюдаемую в ядре клетки. Дальнейшее уплотнение хроматина в ядре клетки необходимо, но это еще не совсем понятно. В настоящее время считается, что повторяющиеся нуклеосомы с промежуточной «линкерной» ДНК образуют 10-нм волокно, описываемое как «бусинки на нити», и имеют коэффициент упаковки примерно от пяти до десяти. Цепочка нуклеосом может быть расположена в волокне 30 нм, компактной структуре с коэффициентом упаковки ~ 50, образование которой зависит от присутствия гистона H1.
Кристаллическая структура тетрануклеосомы была представлена и использована для создания предложенной структуры 30-нм волокна в виде двухстартовой спирали. До сих пор существует определенное количество разногласий по поводу этой модели, поскольку она несовместима с недавними данными электронной микроскопии. Помимо этого, структура хроматина плохо изучена, но классически предполагается, что 30 нм волокна расположены в виде петель вдоль центрального белкового каркаса с образованием транскрипционно активного эухроматина. Дальнейшее уплотнение приводит к транскрипционно неактивному гетерохроматину.
Хотя нуклеосома является очень стабильным комплексом белок-ДНК, она не статична и, как было показано, претерпевает ряд различных структурных перестроек, включая скольжение нуклеосом и экспонирование участков ДНК. В зависимости от контекста нуклеосомы могут ингибировать или облегчать связывание фактора транскрипции. Положения нуклеосом контролируются тремя основными факторами: во-первых, внутренняя аффинность связывания гистонового октамера зависит от последовательности ДНК. Во-вторых, нуклеосома может вытесняться или рекрутироваться за счет конкурентного или кооперативного связывания других белковых факторов. В-третьих, нуклеосома может активно перемещаться с помощью АТФ-зависимых ремоделирующих комплексов.
Работа, выполненная в лаборатории Брэдбери, показала, что нуклеосомы, восстановленные на позиционной последовательности 5S ДНК, были способны репозиционировать себя трансляционно на соседние последовательности при термической инкубации. Более поздняя работа показала, что это репозиционирование не требует разрушения октамера гистонов, но согласуется с тем, что нуклеосомы способны «скользить» по ДНК в цис. В 2008 году было дополнительно обнаружено, что сайты связывания CTCF действуют как якоря позиционирования нуклеосом, так что при использовании для выравнивания различных геномных сигналов можно легко идентифицировать множественные фланкирующие нуклеосомы. Хотя нуклеосомы по своей природе подвижны, эукариоты развили большое семейство АТФ-зависимых ферментов ремоделирования хроматина для изменения структуры хроматина, многие из которых делают это посредством скольжения нуклеосом. В 2012 году лаборатория Бины Пиллаи продемонстрировала, что скольжение нуклеосом является одним из возможных механизмов крупномасштабной тканеспецифической экспрессии генов. Работа показывает, что сайт начала транскрипции для генов, экспрессируемых в конкретной ткани, истощены по нуклеосомам, в то время как тот же набор генов в другой ткани, где они не экспрессируются, связаны с нуклеосомами.
Работа лаборатории Widom показала, что нуклеосомная ДНК находится в равновесии между завернутым и развернутым состояниями. Измерения этих скоростей с использованием FRET с временным разрешением показали, что ДНК в нуклеосоме остается полностью обернутой только в течение 250 мс, после чего она разворачивается в течение 10-50 мс, а затем быстро перематывается. Это означает, что ДНК не нужно активно отделять от нуклеосомы, но есть значительная часть времени, в течение которой она полностью доступна. В самом деле, это можно распространить на наблюдение, что введение ДНК-связывающей последовательности в нуклеосому увеличивает доступность соседних участков ДНК при связывании. Эта склонность ДНК в нуклеосоме «дышать» имеет важные функциональные последствия для всех ДНК-связывающих белков, которые действуют в среде хроматина. В частности, динамическое дыхание нуклеосом играет важную роль в ограничении продвижения РНК-полимеразы II во время удлинения транскрипции.
Промоторы активных генов имеют участки, свободные от нуклеосом (NFR). Это обеспечивает доступность промоторной ДНК для различных белков, таких как факторы транскрипции. Свободная от нуклеосом область обычно охватывает 200 нуклеотидов у S. cerevisae. Хорошо расположенные нуклеосомы образуют границы NFR. Эти нуклеосомы называются + 1-нуклеосомой и -1-нуклеосомой и расположены на канонических расстояниях ниже и выше, соответственно, от сайта начала транскрипции. + 1-нуклеосома и несколько нижестоящих нуклеосом также имеют тенденцию включать вариант гистона H2A.Z.
Геномы эукариот повсеместно связаны с хроматином; однако клетки д. пространственно и временно регулировать специфические локусы независимо от основной массы хроматина. Чтобы достичь высокого уровня контроля, необходимого для координации ядерных процессов, таких как репликация, репарация и транскрипция ДНК, клетки разработали множество средств для локальной и специфической модуляции структуры и функции хроматина. Это может включать ковалентную модификацию гистонов, включение вариантов гистонов и нековалентное ремоделирование с помощью АТФ-зависимых ферментов ремоделирования.
 Хвосты гистонов и их функция в образовании хроматина
Хвосты гистонов и их функция в образовании хроматина С тех пор, как они были открыты в середине 1960-х, было предсказано, что модификации гистонов влияют на транскрипцию. Тот факт, что большинство обнаруженных ранних посттрансляционных модификаций было сосредоточено внутри удлинений хвоста, которые выступают из ядра нуклеосомы, приводит к двум основным теориям относительно механизма модификации гистонов. Первая из теорий предполагала, что они могут влиять на электростатические взаимодействия между хвостами гистонов и ДНК, «ослабляя» структуру хроматина. Позже было высказано предположение, что комбинации этих модификаций могут создавать связывающие эпитопы, с помощью которых можно рекрутировать другие белки. Недавно, учитывая, что было обнаружено больше модификаций в структурированных областях гистонов, было выдвинуто предположение, что эти модификации могут влиять на взаимодействия гистон-ДНК и гистон-гистон в ядре нуклеосомы. Предполагается, что модификации (такие как ацетилирование или фосфорилирование), которые снижают заряд ядра глобулярного гистона, «ослабляют» ассоциацию ядра-ДНК; сила эффекта зависит от расположения модификации в ядре. Было показано, что некоторые модификации коррелируют с подавлением гена; другие, кажется, коррелируют с активацией генов. Общие модификации включают ацетилирование, метилирование, или убиквитинирование из лизина ; метилирование из аргинина ; и фосфорилирование по серину. Информация, хранящаяся таким образом, считается эпигенетической, поскольку она не закодирована в ДНК, но по-прежнему передается дочерним клеткам. Поддержание репрессированного или активированного статуса гена часто необходимо для клеточной дифференциации.
Хотя гистоны в значительной степени консервативны на протяжении всей эволюции, было идентифицировано несколько вариантных форм. Эта диверсификация функции гистонов ограничивается H2A и H3, при этом H2B и H4 в основном инвариантны. H2A может быть заменен H2AZ (что приводит к снижению стабильности нуклеосом) или H2AX (который связан с репарацией ДНК и дифференцировкой Т-клеток ), тогда как неактивные Х-хромосомы у млекопитающих обогащены macroH2A. H3 может быть заменен на H3.3 (который коррелирует с активирующими генами и регуляторными элементами), а в центромерах H3 заменяется на CENPA.
Ряд различных реакций связан с термином АТФ-зависимое ремоделирование хроматина. Было показано, что ремоделирующие ферменты скользят по нуклеосомам вдоль ДНК, нарушают контакты гистонов с ДНК до такой степени, что дестабилизируют димер H2A / H2B и вызывают отрицательное супспиральное скручивание ДНК и хроматина. Недавно было показано, что фермент ремоделирования Swr1 вводит вариантный гистон H2A.Z в нуклеосомы. В настоящее время неясно, представляют ли все они отдельные реакции или просто альтернативные результаты общего механизма. Что общего у всех, и это действительно отличительный признак АТФ-зависимого ремоделирования хроматина, так это то, что все они приводят к изменению доступности ДНК.
Исследования, посвященные активации генов in vivo и, что более удивительно, ремоделированию in vitro, показали, что события ремоделирования хроматина и связывания факторов транскрипции носят циклический и периодический характер. Хотя последствия этого для механизма реакции ремоделирования хроматина неизвестны, динамический характер системы может позволить ей быстрее реагировать на внешние раздражители. Недавнее исследование показывает, что положения нуклеосом значительно меняются во время развития эмбриональных стволовых клеток мыши, и эти изменения связаны со связыванием онтогенетических факторов транскрипции.
Исследования 2007 года каталогизировали положения нуклеосом в дрожжах и показали, что нуклеосомы истощены в промоторных областях и источниках репликации. Около 80% генома дрожжей, по-видимому, покрыто нуклеосомами, и образец расположения нуклеосом явно относится к участкам ДНК, которые регулируют транскрипцию, участкам, которые транскрибируются, и участкам, которые инициируют репликацию ДНК. Совсем недавно новое исследование изучало динамические изменения репозиции нуклеосом во время глобального события транскрипционного репрограммирования, чтобы выяснить влияние на смещение нуклеосом во время полногеномных транскрипционных изменений у дрожжей ( Saccharomyces cerevisiae ). Результаты показали, что нуклеосомы, локализованные в промоторных областях, смещаются в ответ на стресс (например, тепловой шок ). Кроме того, удаление нуклеосом обычно соответствовало активации транскрипции, а замена нуклеосом обычно соответствовала репрессии транскрипции, предположительно потому, что сайты связывания факторов транскрипции стали более или менее доступными, соответственно. В общем, только одна или две нуклеосомы были репозиционированы на промоторе, чтобы вызвать эти транскрипционные изменения. Однако даже в хромосомных областях, которые не были связаны с транскрипционными изменениями, наблюдали репозиционирование нуклеосом, предполагая, что покрытие и раскрытие транскрипционной ДНК не обязательно вызывает событие транскрипции. После транскрипции область рДНК должна быть защищена от любого повреждения, это предполагает, что белки HMGB играют основную роль в защите области, свободной от нуклеосом.
 Схема сборки нуклеосом.
Схема сборки нуклеосом. Нуклеосомы можно собирать in vitro с использованием очищенных нативных или рекомбинантных гистонов. Один из стандартных методов загрузки ДНК вокруг гистонов включает использование солевого диализа. Реакция, состоящая из октамеров гистонов и голой матрицы ДНК, может быть инкубирована вместе при концентрации соли 2 М. Постепенно уменьшая концентрацию соли, ДНК уравновесится до положения, в котором она оборачивается вокруг октамеров гистонов, образуя нуклеосомы. В соответствующих условиях этот процесс восстановления позволяет экспериментально картировать сродство нуклеосомного позиционирования данной последовательности.
Недавний прогресс в производстве ядер нуклеосомных частиц с повышенной стабильностью включает сайт-специфичные дисульфидные поперечные связи. В ядерную частицу нуклеосомы могут быть введены две разные поперечные сшивки. Первый сшивает две копии H2A через введенный цистеин (N38C), в результате чего образуется октамер гистона, который устойчив к потере димера H2A / H2B во время восстановления нуклеосом. Вторая перекрестная связь может быть введена между N-концевым гистоновым хвостом H3 и концами нуклеосомной ДНК через встроенный конвертируемый нуклеотид. Сшивка октамера ДНК-гистона стабилизирует ядерную частицу нуклеосомы против диссоциации ДНК при очень низких концентрациях частиц и при повышенных концентрациях солей.
 Этапы сборки нуклеосом
Этапы сборки нуклеосом Нуклеосомы - это основная единица упаковки ДНК, построенная из гистоновых белков, вокруг которых спиралью ДНК. Они служат каркасом для формирования структуры хроматина более высокого порядка, а также для уровня регуляторного контроля экспрессии генов. Нуклеосомы быстро собираются на вновь синтезированной ДНК позади репликационной вилки.
Гистоны H3 и H4 из разобранных старых нуклеосом хранятся поблизости и случайным образом распределяются на вновь синтезированной ДНК. Они собираются комплексом фактора сборки хроматина-1 (CAF-1), который состоит из трех субъединиц (p150, p60 и p48). Недавно синтезированные H3 и H4 собираются с помощью фактора сборки сцепления репликации (RCAF). RCAF содержит субъединицу Asf1, которая связывается с вновь синтезированными белками H3 и H4. Старые белки H3 и H4 сохраняют свои химические модификации, что способствует передаче эпигенетической сигнатуры. Вновь синтезированные белки H3 и H4 постепенно ацетилируются по различным остаткам лизина как часть процесса созревания хроматина. Также считается, что старые белки H3 и H4 в новых нуклеосомах привлекают модифицирующие гистоны ферменты, которые маркируют новые гистоны, внося свой вклад в эпигенетическую память.
В отличие от старых H3 и H4, старые гистоновые белки H2A и H2B высвобождаются и разрушаются; следовательно, недавно собранные белки H2A и H2B включаются в новые нуклеосомы. H2A и H2B собираются в димеры, которые затем загружаются в нуклеосомы с помощью белка сборки нуклеосом-1 (NAP-1), который также способствует скольжению нуклеосом. Нуклеосомы также разделены АТФ-зависимыми нуклеосомными ремоделирующими комплексами, содержащими ферменты, такие как Isw1, Ino80 и Chd1, и впоследствии собираются в структуру более высокого порядка.


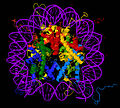
Кристаллическая структура ядерной частицы нуклеосомы ( PDB : 1EQZ ) - различные виды, показывающие детали укладки и организации гистонов. Гистоны H2A, H2B, H3, H4 и ДНК окрашены.