
Войти
Центральная догма молекулярная биология - это объяснение потока генетической информации в биологической системе. Часто говорят, что «ДНК создает РНК, а РНК создает белок», хотя это не его первоначальное значение. Впервые об этом заявил Фрэнсис Крик в 1957 году, а затем опубликовал в 1958 году:
Центральная догма. Это означает, что как только «информация» перешла в белок, она не может выйти снова. Более подробно, передача информации от нуклеиновой кислоты к нуклеиновой кислоте или от нуклеиновой кислоты к белку может быть возможна, но передача от белка к белку или от белка к нуклеиновой кислоте невозможна. Информация означает здесь точное определение последовательности оснований в нуклеиновой кислоте или аминокислотных остатков в белке.
— Francis Crick, 1958и повторно указано в опубликованной статье Nature в 1970 году:
 Информационный поток в биологических системах
Информационный поток в биологических системах Центральная догма молекулярной биологии связана с подробным поэтапным переносом последовательной информации. В нем говорится, что такая информация не может быть передана обратно от белка ни к белку, ни к нуклеиновой кислоте.
— Фрэнсис КрикВторая версия центральной догмы популярна, но неверна. Это упрощенный путь ДНК → РНК → белок, опубликованный Джеймсом Уотсоном в первом издании Молекулярной биологии гена (1965). Версия Уотсона отличается от версии Крика, потому что Уотсон описывает двухэтапный процесс (ДНК → РНК и РНК → белок) как центральную догму. Хотя догма, первоначально заявленная Криком, остается в силе и сегодня, версия Уотсона - нет.
Догма представляет собой основу для понимания передачи последовательности информации между несущими информацию биополимерами, в наиболее распространенном или общем случае, в живых организмах. Существует 3 основных класса таких биополимеров: ДНК и РНК (обе нуклеиновые кислоты) и белок. Между ними может происходить 3 × 3 = 9 возможных прямых передач информации. Догма подразделяет их на 3 группы по 3 человека: три общих переноса (которые, как считается, обычно происходят в большинстве клеток), три особых переноса (известные, что происходят, но только при определенных условиях в случае некоторых вирусов или в лаборатории) и три неизвестных переводы (считается, что никогда не произойдет). Общие передачи описывают нормальный поток биологической информации: ДНК можно скопировать в ДНК (репликация ДНК ), информация ДНК может быть скопирована в мРНК (транскрипция ), и белки могут быть синтезированы с использованием информации в мРНК в качестве матрицы (трансляция ). Специальные переносы описывают: копирование РНК из РНК (репликация РНК ), синтез ДНК с использованием матрицы РНК (обратная транскрипция ) и синтез белков непосредственно из матрицы ДНК без использование мРНК. Неизвестные трансферы описывают: копирование белка из белка, синтез РНК с использованием первичной структуры белка в качестве матрицы и синтез ДНК с использованием первичной структуры белка в качестве матрицы - это не считается естественным.
T биополимеры, которые содержат ДНК, РНК и (поли) пептиды, представляют собой линейные полимеры (то есть: каждый мономер связан максимум с двумя другими мономерами). Последовательность их мономеров эффективно кодирует информацию. Передача информации, описываемая центральной догмой, в идеале является достоверной, детерминированной передачей, при которой последовательность одного биополимера используется в качестве шаблона для создания другого биополимера с последовательностью, которая полностью зависит от последовательности исходного биополимера.
| Общие | Особые | Неизвестно |
|---|---|---|
| ДНК → ДНК | РНК → ДНК | белок → ДНК |
| ДНК → РНК | РНК → РНК | белок → РНК |
| РНК → белок | ДНК → белок | белок → белок |
В том смысле, что репликация ДНК должна происходить, если генетический материал должен быть предоставлен для потомства любой клетки, соматической или репродуктивной, копирование ДНК в ДНК, возможно, является фундаментальным шагом в центральной догме. Сложная группа белков, называемая реплисомой, выполняет репликацию информации от родительской цепи к комплементарной дочерней цепи.
Реплисома включает:
Этот процесс обычно происходит во время S-фазы клеточный цикл.
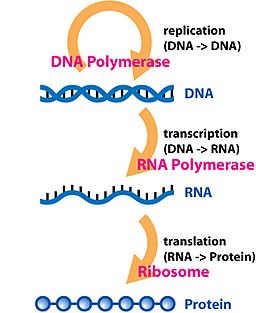
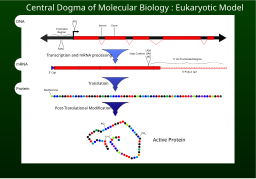
Транскрипция - это процесс, посредством которого информация, содержащаяся в участке ДНК, реплицируется в виде вновь собранной части информационной РНК (мРНК). Ферменты, способствующие этому процессу, включают РНК-полимеразу и факторы транскрипции. В эукариотических клетках первичным транскриптом является пре-мРНК. Пре-мРНК должна быть обработана для продолжения трансляции. Обработка включает добавление 5 'кэпа и поли-A-хвоста к цепи пре-мРНК с последующим сплайсингом. Альтернативный сплайсинг происходит при необходимости, увеличивая разнообразие белков, которые может продуцировать любая отдельная мРНК. Продуктом всего процесса транскрипции (который начался с производства цепи пре-мРНК) является зрелая цепь мРНК.
Зрелая мРНК попадает в рибосому, где она транслируется. В прокариотических клетках, которые не имеют ядерного компартмента, процессы транскрипции и трансляции могут быть связаны вместе без четкого разделения. В эукариотических клетках сайт транскрипции (ядро клетки ) обычно отделен от сайта трансляции (цитоплазма ), поэтому мРНК должна транспортироваться из ядра в цитоплазму, где он может связываться с рибосомами. Рибосома считывает триплет мРНК кодонов, обычно начинающихся с AUG (аденин - урацил - гуанин ) или инициатора кодон метионина ниже сайта связывания рибосомы. Комплексы факторов инициации и факторов элонгации приводят к аминоацилированным транспортным РНК (тРНК) в комплекс рибосома-мРНК, совпадающий с кодоном в мРНК к антикодону на тРНК. Каждая тРНК несет соответствующий остаток аминокислоты для добавления к синтезируемой цепи полипептида. Когда аминокислоты присоединяются к растущей пептидной цепи, цепь начинает складываться в правильную конформацию. Трансляция заканчивается стоп-кодоном , который может быть триплетом UAA, UGA или UAG.
мРНК не содержит всей информации для определения природы зрелого белка. Растущая полипептидная цепь, высвобождаемая из рибосомы, обычно требует дополнительной обработки перед появлением конечного продукта. Во-первых, правильный процесс складывания сложен и жизненно важен. Для большинства белков требуются другие белки-шапероны, чтобы контролировать форму продукта. Некоторые белки затем вырезают внутренние сегменты из собственных пептидных цепей, сращивая свободные концы, которые граничат с разрывом; в таких процессах внутренние «отброшенные» разделы называются интеинами. Другие белки необходимо разделить на несколько частей без сращивания. Некоторые полипептидные цепи необходимо сшить, а другие должны быть присоединены к кофакторам, таким как гем (гем), прежде чем они станут функциональными.
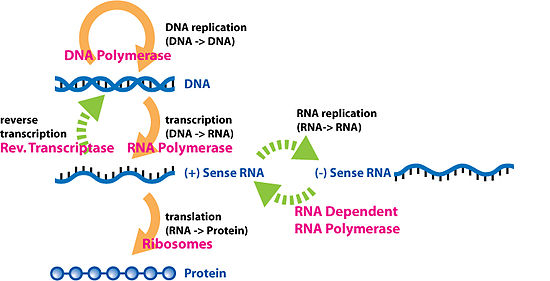 Необычные потоки информации, выделенные зеленым цветом
Необычные потоки информации, выделенные зеленым цветом Обратная транскрипция - это передача информации от РНК к ДНК (обратная нормальной транскрипции). Известно, что это происходит в случае ретровирусов, таких как ВИЧ, а также у эукариот в случае ретротранспозонов и синтез теломер. Это процесс, посредством которого генетическая информация из РНК транскрибируется в новую ДНК.
Репликация РНК - это копирование одной РНК в другую. Так размножаются многие вирусы. Ферменты, копирующие РНК в новую РНК, так называемые РНК-зависимые РНК-полимеразы, также обнаруживаются у многих эукариот, где они участвуют в подавлении РНК.
, в котором последовательность РНК изменяется посредством комплекс белков и «направляющая РНК» также можно рассматривать как перенос РНК на РНК.
Прямая трансляция ДНК в белок была продемонстрирована в бесклеточной системе (т.е. в пробирке) с использованием экстрактов из E. coli, содержащая рибосомы, но не интактные клетки. Эти клеточные фрагменты могут синтезировать белки из одноцепочечных ДНК-матриц, выделенных из других организмов (например, мыши или жабы), и было обнаружено, что неомицин усиливает этот эффект. Однако было неясно, соответствует ли этот механизм трансляции специфическому генетическому коду.
После белка аминокислотные последовательности были переведены из цепей нуклеиновых кислот, они могут редактироваться соответствующими ферментами. Хотя это форма белка, влияющая на последовательность белка, не охватываемая явным образом центральной догмой, существует не так много четких примеров, когда связанные концепции этих двух областей имеют много общего друг с другом.
Интеин - это «паразитарный» сегмент белка, который способен вырезать себя из цепочки аминокислот, когда они выходят из рибосомы, и соединять оставшиеся части с пептидом. связываются таким образом, что основной протеиновый «остов» не распадается. Это случай, когда белок меняет свою собственную первичную последовательность от последовательности, изначально кодированной ДНК гена. Кроме того, большинство интеинов содержат самонаводящуюся эндонуклеазу или домен HEG, который способен обнаруживать копию родительского гена, которая не включает нуклеотидную последовательность интеина. При контакте с копией, свободной от интеина, домен HEG запускает механизм репарации двухцепочечных разрывов ДНК. Этот процесс вызывает копирование последовательности интеина из исходного гена-источника в ген, свободный от интеина. Это пример того, как белок напрямую редактирует последовательность ДНК, а также увеличивает ее наследственное распространение.
Вариация состояний метилирования ДНК может значительно изменить уровни экспрессии гена. Вариации метилирования обычно происходят под действием ДНК метилаз. Если изменение передается по наследству, оно считается эпигенетическим. Когда изменение информационного статуса не передается по наследству, это будет соматический эпитип. Эффективное информационное наполнение было изменено посредством воздействия белка или белков на ДНК, но первичная последовательность ДНК не изменилась.
Прионы представляют собой белки определенных аминокислотных последовательностей в определенных конформациях. Они размножаются в клетках-хозяевах, внося конформационные изменения в другие молекулы белка с той же аминокислотной последовательностью, но с другой конформацией, которая является функционально важной или вредной для организма. Как только белок трансформируется в прионную укладку, он меняет функцию. В свою очередь, он может передавать информацию в новые клетки и реконфигурировать более функциональные молекулы этой последовательности в альтернативную прионную форму. В некоторых типах прионов грибов это изменение является непрерывным и прямым; информационный поток: Белок → Белок.
Некоторые ученые, такие как и Юджин Кунин, утверждали, что наследование, опосредованное прионами, нарушает центральную догму молекулярной биологии. Однако Розалинда Ридли в «Молекулярной патологии прионов» (2001) написала, что «гипотеза прионов не еретична центральной догме молекулярной биологии - информация, необходимая для производства белков, закодирована в нуклеотидной последовательности. нуклеиновой кислоты - потому что он не утверждает, что белки реплицируются. Скорее, он утверждает, что внутри белковых молекул есть источник информации, который способствует их биологической функции, и что эта информация может быть передана другим молекулам ».
Джеймс А. Шапиро утверждает, что надмножество этих примеров следует классифицировать как естественную генную инженерию, и их достаточно, чтобы опровергнуть основную догму. Хотя Шапиро получил уважительное отношение к своей точке зрения, его критики не были убеждены в том, что его понимание центральной догмы соответствует тому, что имел в виду Крик.
В своей автобиографии, What Mad Pursuit Крик написал о своем выборе слова догма и некоторых проблемах, которые оно вызвало у него:
«Я назвал эту идею центральной догмой, я подозреваю, по двум причинам. Я уже использовал очевидное слово гипотеза в гипотезе последовательности, и, кроме того, я хотел предположить, что это новое предположение был более центральным и могущественным... Как выяснилось, использование слова догма доставляло больше хлопот, чем оно того стоило. Много лет спустя Жак Моно указал мне, что я не появлялся понять правильное употребление слова догма, а это убеждение, в котором нельзя сомневаться. Я действительно понимал это смутно, но поскольку я думал, что все религиозные верования не имеют Основание, я использовал это слово так, как я сам думал о нем, а не как большинство людей в мире, а просто применил его к великой гипотезе, которая, как бы ни была правдоподобна, не имела прямого экспериментального подтверждения ».
Точно так же Гораций Фриланд Джадсон записывает в «Восьмой день творения»:
«Я думал, что догма - это идея, для которой не было разумных доказательств. Понимаете ?!» И Крик восторженно взревел. «Я просто не знал, что означают догмы. И с таким же успехом я мог бы назвать это« центральной гипотезой »или - вы знаете. Именно это я хотел сказать.« Догма »была просто крылатой фразой».
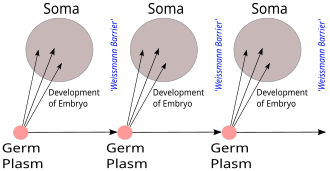 В теории зародышевой плазмы Августа Вейсмана наследственный материал, зародышевая плазма, ограничивается гонадами. Соматические клетки (тела) развиваются заново в каждом поколении из зародышевой плазмы. Что бы ни случилось с этими клетками, это не повлияет на следующее поколение.
В теории зародышевой плазмы Августа Вейсмана наследственный материал, зародышевая плазма, ограничивается гонадами. Соматические клетки (тела) развиваются заново в каждом поколении из зародышевой плазмы. Что бы ни случилось с этими клетками, это не повлияет на следующее поколение. Барьер Вейсмана, предложенный Августом Вейсманном в 1892 году, различает «бессмертные» линии зародышевых клеток (зародышевой плазмы ), которые производят гаметы и «одноразовые» соматические клетки. Наследственная информация перемещается только от клеток зародышевой линии к соматическим клеткам (то есть соматические мутации не наследуются). Это до открытия роли или структуры ДНК не предсказывает основную догму, но предвосхищает ее геноцентрический взгляд на жизнь, хотя и в немолекулярных терминах.
| На Wikimedia Commons есть материалы, связанные с Центральной догмой молекулярной биологии. |