
Войти
| CRISPR-ассоциированная эндонуклеаза Cas9 | |||||||
|---|---|---|---|---|---|---|---|
 Кристаллическая структура S pyogenes Cas9 в комплексе с sgRNA и ее ДНК-мишень с разрешением 2,5. Кристаллическая структура S pyogenes Cas9 в комплексе с sgRNA и ее ДНК-мишень с разрешением 2,5. | |||||||
| Идентификаторы | |||||||
| Организм | Streptococcus pyogenes M1 | ||||||
| Символ | cas9 | ||||||
| Alt. символы | SpCas9 | ||||||
| Entrez | 901176 | ||||||
| PDB | 4OO8 | ||||||
| RefSeq (мРНК) | NC_002737.2 | ||||||
| RefSeq (Prot) | NP_269215.1 | ||||||
| UniProt | Q99ZW2 | ||||||
| Другие данные | |||||||
| Номер EC | 3.1.-.- | ||||||
| Хромосома | Геномный: 0,85 - 0,86 Мб | ||||||
| |||||||
Cas9 (CRISPR как ассоциированный белок 9, ранее называвшийся Cas5, Csn1 или Csx12) представляет собой 160 килодальтон белок, который играет жизненно важную роль в иммунологической защите некоторых бактерий от ДНК-вирусов и плазмид и широко используется в генной инженерии. Приложения. Его основная функция - разрезать ДНК и тем самым изменять геном клетки. Методика редактирования генома CRISPR-Cas9 внесла значительный вклад в Нобелевскую премию по химии в 2020 году, присужденную Эммануэль Шарпантье и Дженнифер Дудна.
С технической точки зрения Cas9 - это двойной РНК -управляемый ДНК эндонуклеаза фермент, связанный с кластерными регулярными короткими палиндромными повторами с регулярными интервалами (CRISPR ) адаптивная иммунная система Streptococcus pyogenes. S. pyogenes использует CRISPR для запоминания и Cas9 для последующего опроса и расщепления чужеродной ДНК, такой как вторгающаяся ДНК бактериофага или плазмидная ДНК. Cas9 выполняет этот запрос, раскручивая чужеродную ДНК и проверяя сайты, комплементарные спейсерной области из 20 пар оснований направляющей РНК. Если ДНК-субстрат комплементарен направляющей РНК, Cas9 расщепляет вторгающуюся ДНК. В этом смысле механизм CRISPR-Cas9 имеет ряд параллелей с механизмом РНК-интерференции (RNAi) у эукариот.
Помимо своей первоначальной функции в бактериальном иммунитете, белок Cas9 широко используется в качестве инструмента геномной инженерии для индуцирования сайт-направленных двухцепочечных разрывов в ДНК. Эти разрывы могут приводить к инактивации генов или введению гетерологичных генов посредством негомологичного соединения концов и гомологичной рекомбинации соответственно во многих лабораторных модельных организмах. Наряду с белками нуклеаз цинкового пальца и эффекторной нуклеазой (Talen), подобной активатору транскрипции (Talen), Cas9 становится важным инструментом в области редактирования генома.
Cas9 приобрел популярность в последние годы, потому что он может расщеплять практически любую последовательность, комплементарную направляющей РНК. Поскольку целевая специфичность Cas9 проистекает из направляющей РНК: комплементарности ДНК, а не из модификаций самого белка (например, TALEN и цинковые пальцы ), создание Cas9 для нацеливания на новую ДНК является простым. Версии Cas9, которые связываются, но не расщепляют родственную ДНК, можно использовать для локализации транскрипционного активатора или репрессоров в конкретных последовательностях ДНК с целью контроля активации и репрессии транскрипции. Для нативного Cas9 требуется направляющая РНК, состоящая из двух разрозненных РНК, которые связываются - РНК CRISPR (crRNA) и трансактивирующей crRNA (tracrRNA ). Нацеливание на Cas9 было упрощено за счет создания химерной одиночной направляющей РНК (chiRNA). Ученые предположили, что основанные на Cas9 генные диски могут быть способны редактировать геномы целых популяций организмов. В 2015 году Cas9 впервые был использован для модификации генома человеческих эмбрионов.
Чтобы выжить во множестве сложных, негостеприимных мест обитания, заполненных бактериофагами, бактериями и архей разработали методы уклонения от хищных вирусов и защиты от них. Сюда входит система адаптивного иммунитета CRISPR. На практике системы CRISPR / Cas действуют как самопрограммируемые рестрикционные ферменты. Локусы CRISPR состоят из коротких палиндромных повторов, которые встречаются через равные промежутки времени, и состоят из альтернативных повторов CRISPR и переменных спейсеров CRISPR длиной 24-48 нуклеотидов. Эти локусы CRISPR обычно сопровождаются соседними генами, ассоциированными с CRISPR (cas). В 2005 году три отдельные группы обнаружили, что спейсерные области гомологичны чужеродным элементам ДНК, включая плазмиды и вирусы. Эти отчеты предоставили первое биологическое свидетельство того, что CRISPR могут функционировать как иммунная система.
Cas9 часто использовался как инструмент редактирования генома. Cas9 был использован в недавних разработках для предотвращения манипулирования вирусами ДНК хозяев. Поскольку CRISPR-Cas9 был разработан на основе систем бактериального генома, его можно использовать для нацеливания на генетический материал вирусов. Использование фермента Cas9 может стать решением многих вирусных инфекций. Cas9 обладает способностью нацеливаться на определенные вирусы путем нацеливания на определенные цепи вирусной генетической информации. Более конкретно, фермент Cas9 нацелен на определенные участки вирусного генома, что мешает вирусу выполнять свои нормальные функции. Cas9 также использовался для разрушения вредных цепей ДНК и РНК, которые вызывают заболевания и мутировавшие цепи ДНК. Cas9 уже показал многообещающие результаты в подавлении эффектов ВИЧ-1. Было показано, что Cas9 подавляет экспрессию длинных концевых повторов в ВИЧ-1. При введении в геном ВИЧ-1 Cas9 показал способность мутировать цепи ВИЧ-1. Cas9 также использовался при лечении гепатита В путем нацеливания на концы некоторых из длинных концевых повторов в геноме вируса гепатита В. Cas9 был использован для исправления мутаций, вызывающих катаракту у мышей.
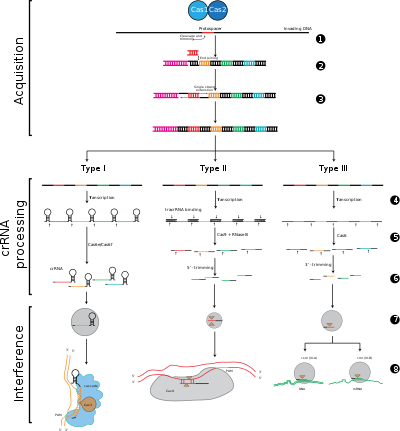 Фиг. 2: Этапы иммунитета к CRISPR
Фиг. 2: Этапы иммунитета к CRISPR Системы CRISPR-Cas подразделяются на три основных типа (тип I, тип II и тип III) и двенадцать подтипов, которые основаны на их генетическом содержании и структурных различиях. Однако основными определяющими характеристиками всех систем CRISPR-Cas являются гены cas и их белки: cas1 и cas2 универсальны для типов и подтипов, в то время как cas3, cas9 и cas10 являются генами-сигнатурами для типа I, тип II и тип III соответственно.
Адаптация включает распознавание и интеграцию спейсеров между двумя соседними повторами в локусе CRISPR. «Протоспейсер» относится к последовательности вирусного генома, которая соответствует спейсеру. Короткий участок консервативных нуклеотидов существует проксимальнее протоспейсера, который называется мотивом, прилегающим к протоспейсеру (PAM). PAM - это мотив узнавания, который используется для получения фрагмента ДНК. При типе II Cas9 распознает PAM во время адаптации, чтобы гарантировать получение функциональных спейсеров.
Экспрессия CRISPR включает транскрипцию первичного транскрипт, называемый CRISPR РНК (пре-crRNA), который транскрибируется из локуса CRISPR с помощью РНК-полимеразы. Затем специфические эндорибонуклеазы расщепляют пре-crRNA на малые CRISPR-РНК (crRNA).
Интерференция включает crRNAs в составе мультибелкового комплекса, называемого CASCADE, которые могут распознавать и специфически соединять пары оснований с участками вставки комплементарной чужеродной ДНК. Комплекс crRNA-чужеродная нуклеиновая кислота затем расщепляется, однако, если есть несоответствия между спейсером и целевой ДНК или если есть мутации в PAM, то расщепление не начнется. В последнем случае чужеродная ДНК не является мишенью для атаки клетки, поэтому репликация вируса продолжается, и хозяин не застрахован от вирусной инфекции. Этап интерференции может механически и временно отличаться от приобретения и выражения CRISPR, но для полноценного функционирования в качестве защитной системы все три фазы должны быть функциональными.
Этап 1: интеграция спейсера CRISPR. Протоспейсеры и связанные с протоспейсерами мотивы (показаны красным) приобретаются на «ведущем» конце массива CRISPR в ДНК хозяина. Массив CRISPR состоит из последовательностей спейсеров (показаны в цветных прямоугольниках), окруженных повторами (черные ромбы). Для этого процесса требуются Cas1 и Cas2 (и Cas9 в типе II), которые закодированы в локусе cas, которые обычно расположены рядом с массивом CRISPR.
Этап 2: выражение CRISPR. Пре-crРНК транскрибируется, начиная с лидерной области, РНК-полимеразой хозяина, а затем расщепляется белками Cas на более мелкие crRNA, содержащие один спейсер и частичный повтор (показаны в виде шпилечной структуры с окрашенными спейсерами).
Этап 3: Вмешательство CRISPR. crRNA со спейсером, который имеет сильную комплементарность с входящей чужеродной ДНК, начинает событие расщепления (изображено ножницами), для которого требуются белки Cas. Расщепление ДНК препятствует репликации вируса и обеспечивает иммунитет хозяину. Этап интерференции может функционально и временно отличаться от получения и выражения CRISPR (обозначен белой линией, разделяющей ячейку).
dCas9, также называемая дефицитной по эндонуклеазе Cas9, может использоваться для редактирования экспрессии гена при применении к сайту связывания транскрипции желаемого участка гена. Оптимальная функция dCas9 объясняется его принципом действия. Экспрессия генов подавляется, когда нуклеотиды больше не добавляются к цепи РНК и, следовательно, прекращают удлинение этой цепи, и в результате влияет на процесс транскрипции. Этот процесс происходит, когда dCas9 производится массово, поэтому он может воздействовать на большинство генов в любой момент времени через специфичную для последовательности молекулу направляющей РНК. Поскольку dCas9, по-видимому, подавляет экспрессию генов, это действие усиливается еще больше, когда его используют в сочетании с репрессивными доменами модификаторов хроматина. Белок dCas9 выполняет другие функции, помимо регуляции экспрессии генов. К белку dCas9 может быть добавлен промотор, который позволяет им работать друг с другом, чтобы стать эффективными при запуске или остановке транскрипции в различных последовательностях вдоль цепи ДНК. Эти два белка специфичны в том, где они действуют на ген. Это преобладает у определенных типов прокариот, когда промотор и dCas9 выравниваются вместе, чтобы препятствовать удлинению полимера нуклеотидов, соединяющихся вместе, с образованием транскрибируемого фрагмента ДНК. Без промотора белок dCas9 не имеет такого же эффекта сам по себе или с телом гена.
При дальнейшем исследовании эффектов репрессии транскрипции, H3K27, аминокислотный компонент гистона, становится метилированным посредством взаимодействие dCas9 и пептида, называемого FOG1. По сути, это взаимодействие вызывает репрессию гена на C + N-концевом участке аминокислотного комплекса в конкретном месте соединения гена и, как следствие, прекращает транскрипцию.
dCas9 также оказывается эффективным, когда это происходит. на изменение определенных белков, которые могут вызвать заболевания. Когда dCas9 присоединяется к форме РНК, называемой направляющей РНК, он предотвращает распространение повторяющихся кодонов и последовательностей ДНК, которые могут быть вредными для генома организма. По сути, когда продуцируются множественные повторяющиеся кодоны, это вызывает ответ или привлекает большое количество dCas9 для борьбы с избыточной продукцией этих кодонов и приводит к остановке транскрипции. dCas9 работает синергетически с гРНК и напрямую влияет на ДНК-полимеразу II, продолжая транскрипцию.
Дальнейшее объяснение того, как работает белок dCas9, можно найти в их использовании геномов растений путем регулирования выработки генов в растениях для увеличения или уменьшения определенных характеристик. Система CRISPR-CAS9 может либо активировать, либо подавлять гены. Белки dCas9 являются компонентом системы CRISPR-CAS9, и эти белки могут репрессировать определенные области гена растения. Это происходит, когда dCAS9 связывается с репрессорными доменами, а в случае растений действительно происходит дезактивация регуляторного гена, такого как AtCSTF64.
Бактерии также являются еще одним направлением использования белков dCas9. Поскольку эукариоты имеют более крупный состав ДНК и геном; с гораздо более мелкими бактериями легко манипулировать. В результате эукариоты используют dCas9 для подавления РНК-полимеразы продолжения процесса транскрипции генетического материала.
 Кристаллическая структура CRISPR-ассоциированного белка Cas9, основанный на PDB 5AXW от Nishimasu et al.
Кристаллическая структура CRISPR-ассоциированного белка Cas9, основанный на PDB 5AXW от Nishimasu et al. Cas9 имеет двухлепестковую архитектуру с направляющей РНК, расположенной между альфа-спиральной долей (синий) и долей нуклеазы (голубой, оранжевый и серый). Эти два лепестка соединены единой мостовой спиралью. Есть два нуклеазных домена, расположенных в доле многодоменной нуклеазы: RuvC (серый), который расщепляет нецелевую цепь ДНК, и нуклеазный домен HNH (голубой), который расщепляет целевую цепь ДНК. Домен RuvC кодируется последовательно разнесенными сайтами, которые взаимодействуют в третичной структуре с образованием домена расщепления RuvC (см. Правый рисунок).
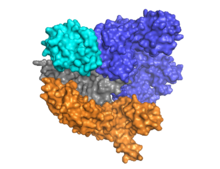 Кристаллическая структура Cas9 в форме Apo. Структурное воспроизведение было выполнено с использованием программного обеспечения UCSF Chimera.
Кристаллическая структура Cas9 в форме Apo. Структурное воспроизведение было выполнено с использованием программного обеспечения UCSF Chimera. Ключевой особенностью ДНК-мишени является то, что она должна содержать мотив, прилегающий к протоспейсер, (PAM), состоящий из трехнуклеотидной последовательности - NGG. Этот PAM распознается взаимодействующим с PAM доменом (домен PI, оранжевый), расположенным рядом с C-концевым концом Cas9. Cas9 претерпевает явные конформационные изменения между состояниями апо, связанными направляющими РНК и связанными направляющими РНК: ДНК.
Cas9 распознает архитектуру стебель-петля, присущую локусу CRISPR, которая опосредует созревание комплекса crRNA-tracrRNA рибонуклеопротеин. Cas9 в комплексе с CRISPR РНК (crRNA) и трансактивирующей crRNA (tracrRNA) дополнительно распознает и разрушает целевую дцДНК. В представленной здесь сокристаллической структуре комплекс crRNA-tracrRNA заменен химерной однонаправленной РНК (sgRNA, выделена красным цветом), которая, как было доказано, выполняет ту же функцию, что и природный комплекс РНК. Основание sgRNA в паре с целевой ssDNA заякорено Cas9 в виде Т-образной архитектуры. Эта кристаллическая структура ДНК-связанного фермента Cas9 показывает отчетливые конформационные изменения в альфа-спиральной доле по отношению к доле нуклеазы, а также расположение домена HNH. Белок состоит из доли узнавания (REC) и доли нуклеазы (NUC). Все области, кроме HNH, образуют тесные взаимодействия друг с другом и комплекс sgRNA-ssDNA, в то время как домен HNH образует мало контактов с остальной частью белка. В другой конформации комплекса Cas9, наблюдаемой в кристалле, домен HNH не виден. Эти структуры предполагают конформационную гибкость домена HNH.
На сегодняшний день изучены и опубликованы как минимум три кристаллические структуры. Один представляет конформацию Cas9 в состоянии апо, а два представляет Cas9 в состоянии, связанном с ДНК.
 CRISPR / Cas9
CRISPR / Cas9 В комплексе sgRNA-Cas9, в зависимости от кристаллической структуры, REC1, BH и PI домены имеют важные контакты с остовом или основаниями как в области повтора, так и в области спейсера. Было протестировано несколько мутантов Cas9, включая делецию доменов REC1 или REC2 и мутации остатков в BH. Мутанты, родственные REC1 и BH, проявляют более низкую активность или ее отсутствие по сравнению с диким типом, что указывает на то, что эти два домена имеют решающее значение для распознавания sgRNA в повторяющейся последовательности и стабилизации всего комплекса. Хотя взаимодействия между последовательностью спейсера и Cas9, а также доменом PI и районом повтора нуждаются в дальнейших исследованиях, сокристалл демонстрирует четкую границу раздела между Cas9 и sgRNA.
Предыдущий анализ последовательности и биохимические исследования показали, что Cas9 содержит два нуклеазных домена: McrA-подобный домен HNH-нуклеазы и RuvC-подобный нуклеазный домен. Эти HNH- и RuvC-подобные нуклеазные домены ответственны за расщепление комплементарной / целевой и некомплементарной / нецелевой цепей ДНК, соответственно. Несмотря на низкое сходство последовательностей, последовательность, подобная РНКазе H, имеет складку RuvC (один член семейства РНКазы H), а область HNH складывается как T4 Endo VII (один член семейства эндонуклеаз HNH).
Дикий тип S. pyogenes Cas9 требует кофакторов магния (Mg) для РНК-опосредованного расщепления ДНК; однако было показано, что Cas9 проявляет различные уровни активности в присутствии ионов других двухвалентных металлов. Например, было показано, что Cas9 в присутствии марганца (Mn) способен к РНК-независимому расщеплению ДНК. кинетика расщепления ДНК Cas9 вызвала большой интерес для научного сообщества, поскольку эти данные позволяют понять тонкости реакции. Хотя было показано, что расщепление ДНК связанным с РНК Cas9 относительно быстрое (k ≥ 700 с), высвобождение продуктов расщепления происходит очень медленно (t 1/2 = ln (2) / k ≈ 43-91 ч), по существу превращая Cas9 в фермент с одним оборотом. Дополнительные исследования кинетики Cas9 показали, что сконструированный Cas9 эффективен в снижении эффектов за счет изменения скорости реакции.
Большинство археи и бактерии упорно отказываются позволить cas9 редактировать их геном. Это потому, что они могут прикреплять чужеродную ДНК, которая не влияет на них, в свой геном. Другой способ, которым эти клетки игнорируют Cas9, - это система процесса рестрикционной модификации (RM). Когда бактериофаг проникает в бактериальную клетку или клетку архей, он становится мишенью для системы RM. Затем система RM разрезает ДНК бактериофагов на отдельные части рестрикционными ферментами и использует эндонуклеазы для дальнейшего разрушения цепей ДНК. Это создает проблему для редактирования Cas9, потому что система RM также нацелена на чужеродные гены, добавленные процессом Cas9.
Из-за уникальной способности Cas9 связываться практически с любой последовательностью комплемента в любом геноме, исследователи хотели использовать этот фермент для подавления транскрипции различных геномных локусов. Для этого два критических каталитических остатка домена RuvC и HNH могут быть мутированы в аланин, устраняя всю эндонуклеазную активность Cas9. Полученный в результате белок, обозначенный как «мертвый» Cas9 или для краткости «dCas9», все еще может прочно связываться с дцДНК. Этот каталитически неактивный вариант Cas9 был использован как для механистических исследований вопросительного связывания Cas9 ДНК, так и в качестве общего программируемого ДНК-связывающего комплекса РНК-белок.
Взаимодействие dCas9 с дцДНК-мишенью настолько тесное, что денатурирующий белок с высокой молярностью мочевина не может полностью диссоциировать комплекс dCas9 РНК-белок от дцДНК-мишени. dCas9 был нацелен с помощью сконструированных единственных направляющих РНК к сайтам инициации транскрипции любых локусов, где dCas9 может конкурировать с РНК-полимеразой на промоторах, останавливая транскрипцию. Кроме того, dCas9 может быть нацелен на кодирующую область локусов, так что ингибирование РНК-полимеразы происходит во время фазы элонгации транскрипции. У эукариот подавление экспрессии генов может быть увеличено путем нацеливания dCas9 на энхансерные последовательности, где dCas9 может блокировать сборку факторов транскрипции, приводя к подавлению экспрессии специфических генов. Более того, направляющие РНК, предоставленные для dCas9, могут быть разработаны таким образом, чтобы включать определенные несовпадения с его комплементарной родственной последовательностью, что количественно ослабит взаимодействие dCas9 с его запрограммированной родственной последовательностью, позволяя исследователю настроить степень подавления гена, применяемого к интересующему гену. Эта технология в принципе аналогична РНКи, так что экспрессия генов модулируется на уровне РНК. Однако подход dCas9 получил большую популярность, так как существует меньше нецелевых эффектов и в целом более крупные и воспроизводимые эффекты сайленсинга за счет использования dCas9 по сравнению с экранами RNAi. Кроме того, поскольку подход dCas9 к подавлению гена может контролироваться количественно, исследователь теперь может точно контролировать степень репрессии интересующего гена, что позволяет получить ответы на больше вопросов о регуляции гена и стехиометрии гена.
Помимо прямого связывания dCas9 с транскрипционно чувствительными положениями локусов, dCas9 может быть слит с множеством модулирующих белковых доменов для выполнения множества функций. Недавно dCas9 был слит с белками ремоделирования хроматина (HDAC / HAT) для реорганизации структуры хроматина вокруг различных локусов. Это важно для нацеливания на различные интересующие эукариотические гены, поскольку структуры гетерохроматина препятствуют связыванию Cas9. Кроме того, поскольку Cas9 может реагировать на гетерохроматин, предполагается, что этот фермент может быть дополнительно применен для изучения структуры хроматина различных локусов. Кроме того, dCas9 использовался в полногеномном скрининге репрессии генов. Используя большие библиотеки направляющих РНК, способных воздействовать на тысячи генов, был проведен генетический скрининг в масштабе всего генома с использованием dCas9.
Другой метод подавления транскрипции с помощью Cas9 - это прямое расщепление продуктов мРНК с помощью каталитически активного фермента Cas9. Этот подход стал возможным благодаря гибридизации оцДНК с комплементарной последовательностью PAM с оцРНК, позволяющей создать сайт дцДНК-РНК PAM для связывания Cas9. Эта технология дает возможность выделять эндогенные РНК-транскрипты в клетках без необходимости индуцировать химические модификации РНК или методов маркировки РНК.
В отличие от генов, подавляющих молчание, dCas9 также может использоваться для активации генов при слиянии с факторами активации транскрипции. Эти факторы включают субъединицы бактериальной РНК-полимеразы II и традиционные факторы транскрипции у эукариот. Недавно были проведены полногеномные скрининги активации транскрипции с использованием слияния dCas9 под названием «CRISPRa» для активации.