Войти
 | |||
| |||
| Имена | |||
|---|---|---|---|
| Название IUPAC Proline | |||
| Систематическое название IUPAC Пирролидин- 2-карбоновая кислота | |||
| Идентификаторы | |||
| Номер CAS | |||
| 3D-модель (JSmol ) | |||
| Справочник Байльштейна | 80812 | ||
| ЧЭБИ | |||
| ChEMBL |
| ||
| ChemSpider | |||
| DrugBank | |||
| ECHA InfoCard | 100.009.264 | ||
| Номер ЕС |
| ||
| Ссылка на Гмелин | 26927 | ||
| KEGG | |||
| MeSH | Proline | ||
| PubChem CID | |||
| Номер RTECS |
| ||
| UNII | |||
| CompTox Панель управления (EPA ) | |||
InChI
| |||
УЛЫБКИ
| |||
| Свойства | |||
| Химическая формула | C5H9NO2 | ||
| Молярная масса | 115,132 г · моль | ||
| Внешний вид | Прозрачные кристаллы | ||
| Температура плавления | от 205 до 228 ° C (от 401 до 442 ° F; 478-501 K) (разлагается) | ||
| Растворимость | 1,5 г / 100 г этанола 19 ° C | ||
| log P | -0,06 | ||
| Кислотность (pK a) | 1,99 (карбоксил), 10,96 (амино) | ||
| Опасности | |||
| Паспорт безопасности | См.: страница данных | ||
| S-фразы (устаревшие) | S22, S24 / 25 | ||
| Дополнительные данные страница | |||
| Структура и. свойства | Показатель преломления (n),. Диэлектрическая постоянная (εr) и т. д. | ||
| Термодинамические. данные | Фазовое поведение. твердое тело – жидкость – газ | ||
| Спектральные данные | UV, IR, ЯМР, MS | ||
| Если не указано иное, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа). | |||
| Ссылки на информационное окно | |||
Proline (символ Pro или P ) представляет собой органическую кислоту, классифицируемую как протеиногенная аминокислота (используется в биосинтезе белков ), хотя она не содержит аминогруппу -NH. 2, а скорее является вторичным амином. Азот вторичного амина находится в протонированной форме NH 2 в биологических условиях, тогда как карбоксильная группа находится в депротонированной -COO форме. «Боковая цепь» от α-углерода соединяется с азотом, образуя пирролидиновую петлю, классифицируя ее как алифатическую аминокислоту. Он не является незаменимым для человека, то есть организм может синтезировать его из незаменимой аминокислоты L-глутамата. Он кодируется всеми кодонами , начиная с CC (CCU, CCC, CCA и CCG).
Пролин является единственной протеиногенной аминокислотой, которая является вторичным амином, поскольку атом азота присоединен к α-углероду цепочкой из трех других атомов углерода.
Пролин был впервые выделен в 1900 г. Ричард Вильштеттер, получивший аминокислоту при изучении N-метилпролина. Через год после Эмиль Фишер опубликовал синтез пролина из фталимидного пропилмалонового эфира. Название пролин происходит от пирролидина, одного из его компонентов.
Пролин биосинтетически происходит из аминокислоты L-глутамата. Глутамат-5-полуальдегид сначала образуется глутамат-5-киназой (АТФ-зависимой) и глутамат-5-полуальдегиддегидрогеназой (которая требует НАДН или НАДФН). Затем он может либо спонтанно циклизоваться с образованием 1-пирролин-5-карбоновой кислоты, которая восстанавливается до пролина с помощью пирролин-5-карбоксилатредуктазы (с использованием NADH или NADPH), либо превращается в орнитин с помощью орнитинаминотрансферазы с последующей циклизацией орнитинциклодезаминазой с образованием пролина.
 Цвиттерионная структура обоих энантиомеров пролина: (S) -пролин (слева) и (R) -пролин
Цвиттерионная структура обоих энантиомеров пролина: (S) -пролин (слева) и (R) -пролин LБыло обнаружено, что -пролин действует как слабый агонист как рецептора глицина, так и обоих NMDA и не-NMDA (AMPA / каинат ) ионотропные рецепторы глутамата. Было высказано предположение, что это потенциальный эндогенный эксайтотоксин. У растений накопление пролина является обычным физиологическим ответом на различные стрессы, но также является частью программы развития (например, пыльца ).
Отличительные циклические структура боковой цепи пролина придает пролину исключительную конформационную жесткость по сравнению с другими аминокислотами. Это также влияет на скорость образования пептидной связи между пролином и другими аминокислотами. Когда пролин связан как амид в пептидной связи, его азот не связывается с любым водородом, что означает, что он не может действовать как донор водородной связи, но может быть акцептором водородной связи.
Образование пептидной связи с входящей Pro-тРНК происходит значительно медленнее, чем с любыми другими тРНК, что является общей характеристикой N-алкиламинокислот. Образование пептидной связи также происходит медленно между входящей тРНК и цепью, оканчивающейся на пролин; с образованием связей пролин-пролин медленнее всех.
Исключительная конформационная жесткость пролина aff влияет на вторичную структуру белков рядом с остатком пролина и может объяснять более высокую распространенность пролина в белках термофильных организмов. Вторичная структура белка может быть описана в терминах двугранных углов φ, ψ и ω остова белка. Циклическая структура боковой цепи пролина фиксирует угол φ примерно на -65 °.
Пролин действует как структурный разрушитель в середине регулярных элементов вторичной структуры, таких как альфа-спирали и бета-версии ; однако пролин обычно обнаруживается в виде первого остатка альфа-спирали, а также в краевых цепях бета-листов. Пролин также обычно содержится в витках (другой вид вторичной структуры) и помогает в образовании бета-витков. Это может объяснить любопытный факт, что пролин обычно подвергается воздействию растворителя, несмотря на то, что он имеет полностью алифатическую боковую цепь.
Множественные пролины и / или гидроксипролины в ряду могут создавать полипролиновую спираль, преобладающую вторичную структуру в коллагене. гидроксилирование пролина пролилгидроксилазой (или другие добавления электроноакцепторных заместителей, таких как фтор ) увеличивает конформационную стабильность коллагена значительно. Следовательно, гидроксилирование пролина является критическим биохимическим процессом для поддержания соединительной ткани высших организмов. Тяжелые заболевания, такие как цинга, могут быть результатом дефектов этого гидроксилирования, например мутации фермента пролилгидроксилазы или отсутствия необходимого кофактора аскорбата (витамина C).
Пептидные связи с пролином и с другими N-замещенными аминокислотами (такими как саркозин ) могут заселять как цис и транс изомеры. Большинство пептидных связей в подавляющем большинстве принимают транс-изомер (обычно 99,9% в ненапряженных условиях), главным образом потому, что амидный водород (транс-изомер) предлагает меньшее стерическое отталкивание предшествующему атому C α, чем следующий C α атом (цис-изомер). Напротив, цис- и транс-изомеры пептидной связи X-Pro (где X представляет собой любую аминокислоту) испытывают стерические конфликты с соседней заменой и имеют гораздо меньшую разницу в энергии. Следовательно, доля пептидных связей X-Pro в цис-изомере в ненапряженных условиях значительно повышена, причем доли цис обычно находятся в диапазоне 3-10%. Однако эти значения зависят от предыдущей аминокислоты, при этом остатки Gly и ароматические остатки дают повышенные доли цис-изомера. Цис-фракции до 40% были идентифицированы для пептидных связей Aromatic-Pro.
С кинетической точки зрения цис-транс-пролин изомеризация - очень медленный процесс, который может препятствовать развитию сворачивание белка путем улавливания одного или нескольких остатков пролина, важных для укладки в ненативный изомер, особенно когда для нативного белка требуется цис-изомер. Это связано с тем, что остатки пролина синтезируются исключительно в рибосоме в виде транс-изомерной формы. Все организмы обладают ферментами пролилизомераза , чтобы катализировать эту изомеризацию, а некоторые бактерии имеют специализированные пролилизомеразы, связанные с рибосомой. Однако не все пролины необходимы для сворачивания, и сворачивание белка может происходить с нормальной скоростью, несмотря на наличие ненативных конформеров многих пептидных связей X-Pro.
Пролин и его производные часто используются в качестве асимметричных катализаторов в реакциях органокатализа пролина. Восстановление CBS и катализируемая пролином альдольная конденсация являются яркими примерами.
В пивоварении белки, богатые пролином, соединяются с полифенолами с образованием мутности (мутности).
L-Пролин является осмозащитным веществом и поэтому используется во многих фармацевтических и биотехнологических целях.
В питательную среду, используемую в культуре растительной ткани, может быть добавлен пролин. Это может увеличить рост, возможно, потому, что помогает растениям переносить стрессы, связанные с культурой тканей. О роли пролина в стрессовой реакции растений см. § Биологическая активность.
Пролин - одна из двух аминокислот, которые не соответствуют типичному графику Рамачандрана вместе с глицином. Из-за образования кольца, связанного с бета-углеродом, углы ψ и φ вокруг пептидной связи имеют меньше допустимых градусов вращения. В результате он часто встречается в «поворотах» белков, поскольку его свободная энтропия (ΔS) не так велика по сравнению с другими аминокислотами, и, таким образом, в свернутой форме по сравнению с развернутой формой изменение энтропии меньше. Кроме того, пролин редко встречается в α- и β-структурах, так как он снижает стабильность таких структур, поскольку α-N его боковой цепи может образовывать только одну азотную связь.
Кроме того, пролин является единственной аминокислотой, которая не образует красно-пурпурный цвет при проявлении распылением с нингидрином для использования в хроматографии. Пролин, напротив, дает оранжево-желтый цвет.
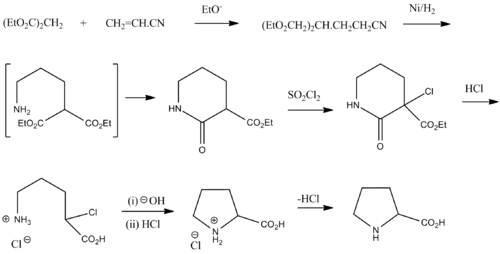
Ричард Вилльштеттер синтезировал пролин реакцией натриевой соли диэтилмалоната с 1,3-дибромпропаном в 1900 г. В 1901 г. Герман Эмиль Фишер выделил пролин из казеина и продуктов разложения γ-фталимидо-пропилмалонового эфира.
рацемический пролин может быть синтезирован из диэтилмалоната и акрилонитрил :