
Войти
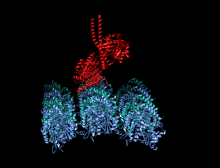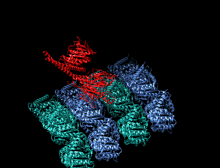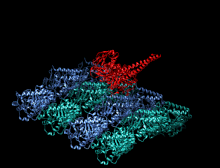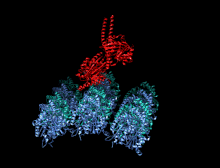 Димер кинезина (красный) прикрепляется к микротрубочкам (синий и зеленый) и перемещается вместе с ними.
Димер кинезина (красный) прикрепляется к микротрубочкам (синий и зеленый) и перемещается вместе с ними.  Анимация «прогулки» кинезина по микротрубочке
Анимация «прогулки» кинезина по микротрубочке A кинезин представляет собой белок, принадлежащий к классу моторных белков, обнаруженных в эукариотических клетках.
Кинезины перемещаются по филаментам микротрубочек (MT) и питаются за счет гидролиза аденозинтрифосфата (АТФ) (таким образом, кинезины являются АТФазами ), тип фермента. Активное движение кинезинов поддерживает несколько клеточных функций, включая митоз, мейоз и транспорт клеточного груза, например, в аксональный транспорт. Большинство кинезинов движется к положительному концу микротрубочек, что в большинстве клеток влечет за собой транспортировку грузов, таких как белковые и мембранные компоненты, от центра клетки к периферии. Этот вид транспорта известен как антероградный транспорт. Напротив, динеины представляют собой моторные белки, которые движутся к минус-концу микротрубочки при ретроградном транспорте.
Кинезины были обнаружены как основанные на МТ двигатели антероградного внутриклеточного транспорта. Член-основатель этого суперсемейства, кинезин-1, был выделен в виде гетеротетрамерного быстрого мотора транспорта аксональных органелл, состоящего из 2 идентичных моторных субъединиц (KHC) и 2 «легких цепей» (KLC), посредством аффинной очистки микротрубочек из экстракты нервных клеток. Впоследствии из экстрактов яиц / эмбрионов иглокожих был очищен другой гетеротримерный мотор на основе МТ, направленный на плюс-конец, названный кинезин-2, состоящий из 2 различных KHC-связанных моторных субъединиц и дополнительной субъединицы «KAP», и наиболее известен благодаря его роль в транспортировке белковых комплексов (частиц IFT) вдоль аксонем во время cilium биогенеза. Молекулярно-генетический и геномный подходы привели к признанию того, что кинезины образуют разнообразное суперсемейство моторов, которые ответственны за множественные события внутриклеточной подвижности в эукариотических клетках. Например, геномы млекопитающих кодируют более 40 белков кинезина, организованных по меньшей мере в 14 семейств, называемых кинезин-1 - кинезин-14.
Члены суперсемейства кинезинов различаются по форме, но прототипный мотор кинезина-1 состоит из двух молекул тяжелой цепи кинезина (KHC), которые образуют белковый димер (пару молекул), который связывает две легкие цепи (KLC), которые уникальны для разных грузов.
Тяжелая цепь кинезина-1 включает глобулярную головку (моторный домен) на аминоконцевом конце, соединенную через короткий гибкий шейный линкер с ножкой - длинный центральный альфа-спиральный домен спиральной спирали, который заканчивается карбоксиконцевым доменом хвостом, который связан с легкими цепями. Стебли двух KHC переплетаются, образуя спиральную катушку , которая направляет димеризацию двух KHC. В большинстве случаев транспортируемый груз связывается с легкими цепями кинезина в последовательности мотива TPR KLC, но в некоторых случаях груз связывается с С-концевыми доменами тяжелых цепей.
| Моторный домен кинезина | |||||||||
|---|---|---|---|---|---|---|---|---|---|
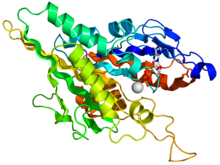 Кристаллографическая структура моторного домена кинезина человека изображена в виде рисунка цвета радуги (N-конец = синий, C-конец = красный) в комплексе с ADP (карандашная диаграмма, углерод = белый, кислород = красный, азот = синий, фосфор = оранжевый) и ионом магния (серая сфера). Кристаллографическая структура моторного домена кинезина человека изображена в виде рисунка цвета радуги (N-конец = синий, C-конец = красный) в комплексе с ADP (карандашная диаграмма, углерод = белый, кислород = красный, азот = синий, фосфор = оранжевый) и ионом магния (серая сфера). | |||||||||
| Идентификаторы | |||||||||
| Symbol | Kinesin motor domain | ||||||||
| Pfam | PF00225 | ||||||||
| InterPro | IPR001752 | ||||||||
| SMART | SM00129 | ||||||||
| PROSITE | PS50067 | ||||||||
| SCOPe | 1bg2 / SUPFAM | ||||||||
| CDD | cd00106 | ||||||||
| |||||||||
Голова является сигнатурой кинезина, и его аминокислотная последовательность хорошо консервативна среди различных кинезинов. Каждая головка имеет два отдельных сайта связывания : один для микротрубочек, а другой для АТФ. Связывание и гидролиз АТФ, а также высвобождение АДФ изменяют конформацию связывающих микротрубочки доменов и ориентацию шейного линкера по отношению к голове; это приводит к движению кинезина. Несколько структурных элементов в Head, включая центральный домен бета-листа и домены Switch I и II, были задействованы как опосредующие взаимодействия между двумя сайтами связывания и шейным доменом. Кинезины структурно связаны с G-белками, которые гидролизуют GTP вместо АТФ. Несколько структурных элементов являются общими для двух семейств, в частности, домена Switch I и Switch II.
 Подвижные и самоингибирующиеся конформации кинезина-1. Самоингибирующаяся конформация: IAK-область хвоста (зеленый) связывается с моторными доменами (желтым и оранжевым), чтобы ингибировать ферментативный цикл кинезина-1. Подвижная конформация: Отсутствие хвоста связываясь, моторные домены кинезина-1 (желтый и оранжевый) могут свободно перемещаться вдоль микротрубочки (MT). PDB 2Y65; PDB 2Y5W.
Подвижные и самоингибирующиеся конформации кинезина-1. Самоингибирующаяся конформация: IAK-область хвоста (зеленый) связывается с моторными доменами (желтым и оранжевым), чтобы ингибировать ферментативный цикл кинезина-1. Подвижная конформация: Отсутствие хвоста связываясь, моторные домены кинезина-1 (желтый и оранжевый) могут свободно перемещаться вдоль микротрубочки (MT). PDB 2Y65; PDB 2Y5W. 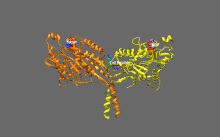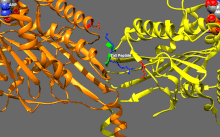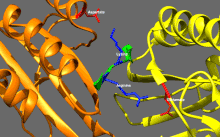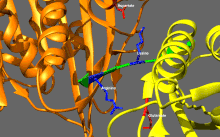 Детальный вид самоингибирования кинезина-1 (показана одна из двух возможных конформаций). Выделение: положительно заряженные остатки (синий) области IAK взаимодействуют в нескольких местах с отрицательно заряженными остатками (красный) моторных доменов PDB 2Y65
Детальный вид самоингибирования кинезина-1 (показана одна из двух возможных конформаций). Выделение: положительно заряженные остатки (синий) области IAK взаимодействуют в нескольких местах с отрицательно заряженными остатками (красный) моторных доменов PDB 2Y65 Кинезины, как правило, имеют низкую базальную ферментативную активность, которая становится значительным при активации микротрубочек. Кроме того, многие члены суперсемейства кинезинов могут самоингибироваться за счет связывания хвостового домена с моторным доменом. Такое самоторможение затем можно устранить с помощью дополнительных правил, таких как привязка к грузу или грузовым адаптерам.
В клетке небольшие молекулы, такие как газы и глюкоза, диффундируют туда, где они необходимы. Большие молекулы, синтезируемые в теле клетки, внутриклеточные компоненты, такие как везикулы и органеллы, такие как митохондрии, слишком велики (а цитозоль слишком переполнен), чтобы иметь возможность распространяются по назначению. Моторные белки выполняют роль транспортировки больших грузов по клетке к нужному месту назначения. Кинезины представляют собой моторные белки, которые транспортируют такой груз, проходя в одном направлении по трекам микротрубочек, гидролизуя одну молекулу аденозинтрифосфата (АТФ) на каждом этапе. Считалось, что гидролиз АТФ приводит в действие каждый шаг, и высвобождаемая энергия продвигает голову вперед к следующему сайту связывания. Однако было высказано предположение, что головка диффундирует вперед, и сила связывания с микротрубочкой - это то, что тянет за собой груз. Кроме того, вирусы, например ВИЧ, используют кинезины, чтобы позволить вирусным частицам перемещаться после сборки.
Имеются убедительные доказательства того, что грузы in-vivo переносятся несколькими двигателями.
Моторные белки перемещаются в определенном направлении по микротрубочке. Микротрубочки полярны; Это означает, что головки связываются с микротрубочкой только в одной ориентации, в то время как связывание АТФ дает каждому шагу свое направление посредством процесса, известного как застегивание шейного линкера.
Ранее было известно, что кинезин перемещает груз в сторону плюса (+) конец микротрубочек, также известный как антероградный транспорт / ортоградный транспорт. Однако недавно было обнаружено, что в почкующихся дрожжевых клетках кинезин Cin8 (член семейства Kinesin-5) также может двигаться в направлении минус-конца или ретроградного транспорта. Это означает, что эти уникальные дрожжевые кинезиновые гомотетрамеры обладают новой способностью двигаться в двух направлениях. До сих пор было показано, что кинезин движется к минус-концу только в группе, при этом двигатели скользят в антипараллельном направлении в попытке разделить микротрубочки. Эта двойная направленность наблюдалась в идентичных условиях, когда свободные молекулы Cin8 движутся к минус-концу, а поперечно-сшивающий Cin8 перемещается к плюс-концам каждой поперечно-сшитой микротрубочки. Одно конкретное исследование проверило скорость, с которой двигались двигатели Cin8, их результаты показали диапазон примерно 25-55 нм / с в направлении полюсов шпинделя. На индивидуальной основе было обнаружено, что при изменении ионных условий двигатели Cin8 могут развивать скорость до 380 нм / с. Предполагается, что двунаправленность дрожжевых моторов кинезина-5, таких как Cin8 и Cut7, является результатом связывания с другими моторами Cin8 и помогает выполнять роль динеина в почкующихся дрожжах, в отличие от человеческого гомолога этих моторов, плюс направил Eg5. Это открытие в белках семейства кинезина-14 (таких как Drosophila melanogaster NCD, почкующиеся дрожжи KAR3 и Arabidopsis thaliana ATK5) позволяет кинезину двигаться в противоположном направлении, к минус концу микротрубочек. Это не типично для кинезина, скорее, исключение из нормального направления движения.
 Диаграмма, иллюстрирующая подвижность кинезина.
Диаграмма, иллюстрирующая подвижность кинезина. Другой тип моторного белка, известный как динеины, движется к минус-концу микротрубочки. Таким образом, они транспортируют груз от периферии ячейки к центру. Примером этого может быть транспорт, происходящий от концевых бутонов нейронального аксона к телу клетки (соме). Это известно как ретроградный транспорт.
Кинезин осуществляет транспортировку путем «ходьбы» по микротрубочке. Было предложено два механизма для объяснения этого движения.
Несмотря на некоторые сохраняющиеся противоречия, все больше экспериментальных данных указывает на то, что механизм передачи рук в руки более вероятен.
Связывание и гидролиз АТФ заставляют кинезин перемещаться по "качелям" механизм »около точки поворота. Этот механизм качелей объясняет наблюдения, что связывание АТФ с безнуклеотидным состоянием, связанным с микротрубочками, приводит к наклону моторного домена кинезина относительно микротрубочки. Критически важно, что до этого наклона шейный линкер не может принять свою состыкованную с моторной головкой форму, обращенную вперед. АТФ-индуцированный наклон дает возможность шейному линкеру стыковаться в этой обращенной вперед конформации. Эта модель основана на моделях CRYO-EM структуры кинезина, связанной с микротрубочками, которые представляют начальное и конечное состояния процесса, но не могут разрешить точные детали перехода между структурами.
Был предложен ряд теоретических моделей кинезина молекулярного моторного белка. Многие проблемы встречаются в теоретических исследованиях, учитывая остающуюся неопределенность в отношении роли белковых структур, точного способа преобразования энергии АТФ в механическую работу и роли тепловых флуктуаций. Это довольно активное направление исследований. Особенно необходимы подходы, которые лучше увязывают с молекулярной архитектурой белка и данными, полученными в результате экспериментальных исследований.
Динамика одной молекулы уже хорошо описана, но кажется, что эти наномасштабные машины обычно работают в больших группах. Недавние экспериментальные исследования показали, что кинезины, перемещаясь по микротрубочкам, взаимодействуют друг с другом, причем взаимодействия являются короткодействующими и слабыми (1,6 ± 0,5 K B T). Одна из разработанных моделей учитывает эти взаимодействия частиц, причем динамические скорости меняются в соответствии с энергией взаимодействия. Если энергия положительна, скорость создания связей (q) будет выше, а скорость разрыва связей (r) будет ниже. Можно понять, что скорость входа и выхода из микротрубочек также будет изменяться под действием энергии (см. Рисунок 1 в ссылке 30). Если второй сайт занят, скорость входа будет α * q, а если предпоследний сайт занят, скорость выхода будет β * r. Этот теоретический подход согласуется с результатами моделирования Монте-Карло для этой модели, особенно для предельного случая очень большой отрицательной энергии. Обычный полностью асимметричный простой процесс исключения для результатов (или TASEP) может быть восстановлен из этой модели, сделав энергию равной нулю.

В последние годы было обнаружено, что молекулярные моторы на основе микротрубочек (включая ряд кинезинов) играют роль в митозе (делении клетки). Кинезины важны для правильной длины веретена и участвуют в раздвижении микротрубочек внутри веретена во время прометафазы и метафазы, а также в деполимеризации минус-концов микротрубочек на центросомах во время анафазы. В частности, белки семейства кинезина-5 действуют внутри веретена, раздвигая микротрубочки, в то время как семейство кинезин 13 действует, деполимеризуя микротрубочки.
Члены суперсемейства кинезинов человека включают следующие белки, которые в стандартизованной номенклатуре, разработанной сообществом исследователей кинезина, разделены на 14 семейств, называемых от кинезин-1 до кинезин-14 :
легкие цепи кинезина-1:
кинезин- 2, ассоциированный белок: