
Войти
Слитые белки или химерные (kī-ˈmir-ik) белки (буквально, состоящие из частей из разных источников) - это белки, созданные путем соединения двух или более генов, которые изначально кодировали отдельные белки. Трансляция этого гибридного гена приводит к одному или нескольким полипептидам с функциональными свойствами, полученными из каждого из исходных белков. Рекомбинантные слитые белки создаются искусственно с помощью технологии рекомбинантных ДНК для использования в биологических исследованиях или терапии. Химеры или химеры обычно обозначают гибридные белки, состоящие из полипептидов, имеющих разные функции или физико-химические структуры. Химерные мутантные белки возникают в природе, когда сложная мутация, такая как хромосомная транслокация, тандемная дупликация или ретротранспозиция, создает новую кодирующую последовательность, содержащую части кодирующих последовательностей из двух разных генов. Встречающиеся в природе слитые белки обычно обнаруживаются в раковых клетках, где они могут функционировать как онкобелки. BCR-ABL слитый белок представляет собой хорошо известный пример онкогенного слитого белка, и считается основным фактором онкогенными хронического миелолейкоза.
Некоторые гибридные белки объединяют целые пептиды и, следовательно, содержат все функциональные домены исходных белков. Однако другие слитые белки, особенно те, которые встречаются в природе, объединяют только части кодирующих последовательностей и, следовательно, не поддерживают исходные функции родительских генов, которые их сформировали.
Многие слияния целых генов полностью функциональны и могут заменять исходные пептиды. Некоторые, однако, испытывают взаимодействия между двумя белками, которые могут изменять их функции. Помимо этих эффектов, некоторые слияния генов могут вызывать регуляторные изменения, которые изменяют, когда и где действуют эти гены. Для частичного слияния генов перетасовка различных активных сайтов и связывающих доменов может привести к появлению новых белков с новыми функциями.
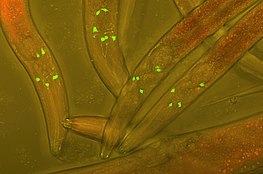 Зеленый флуоресцентный белок (GFP) вставлен в нейроны червей Varbuss для отслеживания развития нейронов.
Зеленый флуоресцентный белок (GFP) вставлен в нейроны червей Varbuss для отслеживания развития нейронов. Слияние флуоресцентных меток с белками в клетке-хозяине - широко популярный метод, используемый в экспериментальных клеточных и биологических исследованиях для отслеживания взаимодействия белков в режиме реального времени. Первая флуоресцентная метка, зеленый флуоресцентный белок (GFP), была выделена из Aequorea Victoria и до сих пор часто используется в современных исследованиях. Более поздние производные включают фотопреобразованные флуоресцентные белки (PCFP), которые впервые были выделены из Anthozoa. Наиболее часто используемым PCFP является флуоресцентная метка Kaede, но разработка зеленого-красного Kikume (KikGR) в 2005 году предлагает более яркий сигнал и более эффективное фотопреобразование. Преимущество использования флуоресцентных меток PCFP заключается в возможности отслеживать взаимодействие перекрывающихся биохимических путей в режиме реального времени. Метка изменит цвет с зеленого на красный, как только белок достигнет интересующей точки пути, и за другим окрашенным белком можно будет следить на протяжении всего пути. Этот метод особенно полезен при изучении путей рециклинга рецепторов, связанных с G-белком (GPCR). Судьбы рециклированных рецепторов G-белка могут быть отправлены на плазматическую мембрану для повторного использования, отмеченную зеленой флуоресцентной меткой, или могут быть отправлены в лизосому для разложения, отмеченную красной флуоресцентной меткой.
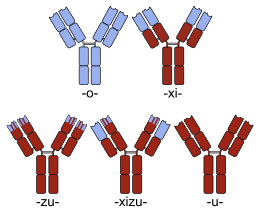 Эскизы мышиных (вверху слева), химерных (вверху справа) и гуманизированных (внизу слева) моноклональных антител. Человеческие части показаны коричневым, а нечеловеческие - синим.
Эскизы мышиных (вверху слева), химерных (вверху справа) и гуманизированных (внизу слева) моноклональных антител. Человеческие части показаны коричневым, а нечеловеческие - синим. Целью создания гибридных белков при разработке лекарств является придание свойств каждого из «родительских» белков полученному химерному белку. В настоящее время для медицинского применения доступны несколько химерных белковых препаратов.
Многие химерные белковые препараты представляют собой моноклональные антитела, специфичность которых в отношении целевой молекулы была разработана с использованием мышей и, следовательно, изначально были «мышиными» антителами. Будучи нечеловеческими белками, мышиные антитела, как правило, вызывают иммунную реакцию при введении человеку. Процесс химеризации включает конструирование замены сегментов молекулы антитела, которые отличают ее от человеческого антитела. Например, могут быть введены человеческие константные домены, тем самым устраняя большинство потенциально иммуногенных частей лекарственного средства без изменения его специфичности в отношении намеченной терапевтической мишени. Номенклатура антител указывает этот тип модификации путем вставки -xi- в непатентованное название (например, abci- xi -mab ). Если части вариабельных доменов также заменяются человеческими частями, получают гуманизированные антитела. Хотя концептуально он не отличается от химер, этот тип обозначается с помощью -zu-, например, в dacli- zu -mab. Дополнительные примеры см. В списке моноклональных антител.
Помимо химерных и гуманизированных антител, существуют другие фармацевтические цели для создания химерных конструкций. Этанерцепт, например, представляет собой TNF - alpha блокатор создается с помощью комбинации фактора некроза опухоли рецепторов (TNFR) с иммуноглобулином G 1 Fc сегментом. TNFR обеспечивает специфичность в отношении мишени для лекарственного средства, а Fc-сегмент антитела, как полагают, увеличивает стабильность и доставляемость лекарственного средства. Дополнительные химерные белки, используемые для терапевтического применения, включают:
 Слияние двух генов (BCR-ABL) для кодирования рекомбинантного онкогенного белка.
Слияние двух генов (BCR-ABL) для кодирования рекомбинантного онкогенного белка. Рекомбинантный слитый белок представляет собой белка, созданные с помощью генной инженерии гена слияния. Обычно это включает удаление стоп- кодона из последовательности кДНК, кодирующей первый белок, затем добавление последовательности кДНК второго белка в рамку считывания посредством лигирования или ПЦР с удлинением перекрытия. Эта ДНК - последовательность, затем будет выражена с помощью ячейки в качестве одного белка. Белок может быть сконструирован таким образом, чтобы включать полную последовательность обоих исходных белков или только часть каждого из них.
Если эти два объекта являются белками, часто также добавляются линкерные (или «спейсерные») пептиды, что повышает вероятность того, что белки сворачиваются независимо и ведут себя так, как ожидалось. Особенно в случае, когда линкеры позволяют очищать белок, линкеры в слияниях белков или пептидов иногда конструируют с сайтами расщепления для протеаз или химических агентов, которые обеспечивают высвобождение двух отдельных белков. Этот метод часто используется для идентификации и очистки белков путем слияния белка GST, пептида FLAG или гекса- His- пептида (6xHis-tag), который можно выделить с помощью аффинной хроматографии с никелевой или кобальтовой смолой. Ди- или мультимерные химерные белки могут быть произведены с помощью генной инженерии путем слияния с исходными белками пептидных доменов, которые индуцируют искусственную ди- или мультимеризацию белков (например, стрептавидин или лейциновые застежки-молнии ). Слитые белки также могут быть произведены с присоединенными к ним токсинами или антителами для изучения развития болезни. Промотор гидрогеназы, P SH, был изучен путем конструирования слияния промотор P SH и gfp с использованием репортерного гена зеленого флуоресцентного белка ( gfp).
Новые рекомбинантные технологии позволили улучшить дизайн слитых белков для использования в таких разнообразных областях, как биодетекция, бумажная и пищевая промышленность, а также биофармацевтические препараты. Недавние усовершенствования включают слияние отдельных пептидов или фрагментов белка с областями существующих белков, такими как N- и C-концы, и, как известно, усиливают следующие свойства:
Самые ранние применения дизайна рекомбинантных белков могут быть задокументированы при использовании одиночных пептидных меток для очистки белков в аффинной хроматографии. С тех пор было разработано множество методов конструирования слитых белков для таких разнообразных приложений, как флуоресцентные белковые метки для рекомбинантных лекарственных препаратов слитых белков. Три часто используемых метода конструирования включают тандемное слияние, вставку домена и посттрансляционную конъюгацию.
Представляющие интерес белки просто соединяются встык путем слияния N- или C-концов между белками. Это обеспечивает гибкую перемычку, обеспечивающую достаточное пространство между партнерами по сварке для обеспечения надлежащего складывания. Однако N- или C-концы пептида часто являются решающими компонентами в получении желаемой картины укладки рекомбинантного белка, что делает простое сквозное соединение доменов неэффективным в этом случае. По этой причине часто требуется белковый линкер для поддержания функциональности интересующих белковых доменов.
Этот метод включает слияние последовательных белковых доменов путем кодирования желаемых структур в единую полипептидную цепь, но иногда может потребоваться встраивание домена в другой домен. Этот метод обычно считается более сложным для выполнения, чем тандемное слияние, из-за трудности поиска подходящего сайта лигирования в интересующем гене.
Этот метод сливает белковые домены после рибосомной трансляции интересующих белков, в отличие от генетического слияния до трансляции, используемого в других рекомбинантных технологиях.
 Белок, используемый в качестве линкера в дизайне слитых белков.
Белок, используемый в качестве линкера в дизайне слитых белков. Белковые линкеры способствуют конструированию слитых белков, обеспечивая соответствующее расстояние между доменами, поддерживая правильную укладку белка в том случае, если взаимодействия N или C-концов имеют решающее значение для укладки. Обычно белковые линкеры допускают важные доменные взаимодействия, усиливают стабильность и уменьшают стерические затруднения, что делает их предпочтительными для использования в дизайне слитых белков, даже когда N- и C-концы могут быть слиты. Три основных типа линкеров - гибкие, жесткие и расщепляемые in vivo.
Встречающиеся в природе гены слияния чаще всего создаются, когда хромосомная транслокация заменяет концевые экзоны одного гена интактными экзонами второго гена. Это создает единственный ген, который можно транскрибировать, сплайсировать и транслировать для получения функционального слитого белка. Многие важные онкогены, способствующие развитию рака, представляют собой гибридные гены, полученные таким образом.
Примеры включают:
Антитела представляют собой слитые белки, полученные с помощью V (D) J рекомбинации.
Есть также редкие примеры встречающихся в природе полипептидов, которые кажутся слиянием двух четко определенных модулей, в которых каждый модуль демонстрирует свою характерную активность или функцию независимо от другого. Двумя основными примерами являются: двойная химера PP2C в Plasmodium falciparum (малярийный паразит), в котором каждый модуль PP2C проявляет ферментативную активность протеинфосфатазы 2C, и иммунофилины двойного семейства, которые встречаются в ряде одноклеточных организмов (таких как простейшие паразиты и флавобактерии).) и содержат полноразмерные шаперонные модули циклофилина и FKBP. Эволюционное происхождение такой химеры остается неясным.
| Библиотечные ресурсы о Fusion Protein |