
Войти
| Юрский период | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 201,3 ± 0,2 - ~ 145,0 млн лет PreꞒ Ꞓ О S D C п Т J K Стр. N | |||||||||
| Хронология | |||||||||
| |||||||||
| Этимология | |||||||||
| Формальность имени | Формальный | ||||||||
| Информация об использовании | |||||||||
| Региональное использование | Глобальный ( ICS ) | ||||||||
| Используемая шкала времени | Шкала времени ICS | ||||||||
| Определение | |||||||||
| Хронологическая единица | Период | ||||||||
| Стратиграфическая единица | Система | ||||||||
| Формальность временного интервала | Формальный | ||||||||
| Определение нижней границы | Первое появление аммонита Psiloceras spelae tirolicum. | ||||||||
| Нижняя граница ГССП | Разрез Кухйох, горы Карвендель, Северные известняковые Альпы, Австрия 47.4839 ° N 11.5306 ° E 47 ° 29′02 ″ с.ш., 11 ° 31′50 ″ в.д. / / 47.4839; 11,5306 | ||||||||
| ГССП ратифицирован | 2010 г. | ||||||||
| Определение верхней границы | Формально не определен | ||||||||
| Кандидаты в определение верхней границы |
| ||||||||
| Верхняя граница раздела (-ов) кандидата GSSP | Никто | ||||||||
Юры ( / dʒ ʊ г æ ы. С ɪ к / juu- RASS -ik ) представляет собой геологический период и стратиграфическая система, которая простиралась от конца триасового периода 201.3 миллионов лет назад (Mya) к началу мела Период, примерно 145 млн лет назад. Юрский период представляет собой средний период мезозойской эры и назван в честь Юрских гор, где впервые были обнаружены пласты известняка этого периода.
Начало юры ознаменовалось крупным триасово-юрским вымиранием, связанным с извержением Магматической провинции Центральной Атлантики. Начало тоарского этап начался около 183 миллионов лет назад, и отмечен событием вымирания, связанного с широко распространенной океанической аноксии, подкисление океана, и повышенные температуры, вероятно, вызванные прорезывания Кару-Ferrar крупных магматических провинций. Конец юры, однако, не имеет четкой границы с меловым периодом и является единственной границей между геологическими периодами, которая формально остается неопределенной.
К началу юрского периода суперконтинент Пангея начал раскол на два массива суши: Лавразию на севере и Гондвану на юге. Климат юрского периода был теплее настоящего, и ледяных шапок не было. Леса росли близко к полюсам, с большими засушливыми территориями в более низких широтах.
На суше фауна перешла от триасовой фауны, в которой доминировали динозавроморфы и псевдозухи- архозавры, к фауне, в которой доминировали только динозавры. Первые птицы появились в юрском периоде, эволюционировав из ветви динозавров- теропод. Другие важные события включают появление самых ранних ящериц и эволюцию терианских млекопитающих. Крокодиломорфы совершили переход от наземной к водной жизни. Океаны населяли морские рептилии, такие как ихтиозавры и плезиозавры, в то время как птерозавры были доминирующими летающими позвоночными.
 Портрет Александра Бронгниара, придумавшего термин «юрский период».
Портрет Александра Бронгниара, придумавшего термин «юрский период». Хроностратиграфическая термин «Jurassic» связан с гор Юра, в пределах горной основном после границы Франции и Швейцарии. Название «Джура» происходит от кельтского корня * jor через Gaulish * iuris «лесная гора», которое, заимствованное на латыни как название места, превратилось в Juria и, наконец, в Jura.
Во время поездки по региону в 1795 году немецкий естествоиспытатель Александр фон Гумбольдт определил, что карбонатные отложения в горах Юра геологически отличаются от мушелкалка триасового возраста в Южной Германии, хотя он ошибочно заключил, что они были старше. Затем в 1799 году он назвал их Джура-Калькштейн («Юрский известняк»).
Тридцать лет спустя, в 1829 году, французский натуралист Александр Бронниар опубликовал книгу под названием « Описание ландшафтов, составляющих земную кору», или «Очерк строения известных земель Земли». В этой книге Бронгниарт использовал фразу terrains jurassiques при сопоставлении «Юра-Калькштейна» Гумбольдта с оолитовыми известняками аналогичного возраста в Британии, таким образом придумав и опубликовав термин «юрский период».
Немецкий геолог Леопольд фон Бух в 1839 году установил тройное деление юрского периода, первоначально названное от самого старого до самого молодого: черная юра, коричневая юра и белая юра. Термин « лиас » ранее использовался Уильямом Конибером и Уильямом Филлипсом в 1822 году для обозначения слоев, эквивалентных по возрасту черному юрскому периоду в Англии.
Французский палеонтолог Альсид д'Орбиньи в своих работах между 1842 и 1852 годами разделил юрский период на десять стадий на основе аммонитов и других скоплений окаменелостей в Англии и Франции, семь из которых все еще используются, хотя ни одна из них не сохранила первоначального определения. Немецкий геолог и палеонтолог Фридрих Август фон Кенштедт в 1858 году разделил три серии фон Буха в Швабской Юре на шесть подразделений, определенных по аммонитам и другим окаменелостям.
Немецкий палеонтолог Альберт Оппель в своих исследованиях между 1856 и 1858 годами изменил первоначальную схему д'Орбиньи и далее разделил ярусы на биостратиграфические зоны, основанные в основном на аммонитах. Большинство современных этапов юрского периода были формализованы на Коллоке Юрского периода в Люксембурге в 1962 году.
Юрский период делится на три эпохи : раннюю, среднюю и позднюю. Точно так же в стратиграфии юра делится на нижнеюрскую, среднеюрскую и верхнеюрскую серии. Геологи делят породы юрского периода на стратиграфический набор единиц, называемых стадиями, каждая из которых сформировалась в течение соответствующих временных интервалов, называемых возрастами.
Этапы могут быть определены глобально или регионально. Для глобальной стратиграфической корреляции Международная комиссия по стратиграфии (ICS) ратифицирует глобальные ярусы на основе разреза и точки глобальной границы (GSSP) из одной формации ( стратотипа ), определяющей нижнюю границу яруса. Возраст юрского периода от самого младшего до самого старшего следующий:
| Серия / эпоха | Стадия / возраст | Нижняя граница |
| Раннемеловой период | Берриасский | ~ 145 млн лет назад |
| Верхняя / Поздняя юра | Титонский | 152,1 ± 0,9 млн лет назад |
| Кимериджский | 157,3 ± 1,0 млн лет назад | |
| Оксфордский | 163,5 ± 1,0 млн лет назад | |
| Средняя юра | Келловейский | 166,1 ± 1,2 млн лет назад |
| Батонский | 168,3 ± 1,3 млн лет назад | |
| Байосский | 170,3 ± 1,4 млн лет назад | |
| Ааленский | 174,1 ± 1,0 млн лет назад | |
| Нижняя / ранняя юра | Тоарский | 182,7 ± 0,7 млн лет назад |
| Плинсбахский | 190,8 ± 1,0 млн лет назад | |
| Синемурийский | 199,3 ± 0,3 млн лет назад | |
| Геттангский | 201,3 ± 0,2 млн лет назад |
 Сложенные нижнеюрские известняковые пласты покрова Долденхорн в Гастеретале, Швейцария
Сложенные нижнеюрские известняковые пласты покрова Долденхорн в Гастеретале, Швейцария  Отложения средней юры в провинции Неукен, Аргентина
Отложения средней юры в провинции Неукен, Аргентина  Тидвелл, член формации Моррисон (верхняя юра), Колорадо
Тидвелл, член формации Моррисон (верхняя юра), Колорадо Стратиграфия юрского периода в первую очередь основана на использовании аммонитов в качестве индексных окаменелостей. Первое появление данное специфических аммонитовых таксонов используются для обозначения зачатков этапов, а также меньшие timespans в пределах этапов, называемых «аммонитовые зонами»; они, в свою очередь, также иногда подразделяются на подзоны. Глобальная стратиграфия основана на стандартных европейских аммонитовых зонах, а другие регионы откалиброваны по европейским сукцессиям.
 База Aalenian GSSP в Фуэнтельсазе
База Aalenian GSSP в Фуэнтельсазе Верхняя граница юрского периода в настоящее время не определена, а граница юрско-мелового периода в настоящее время является единственной границей системы, на которой отсутствует определенный GSSP. Размещение GSSP для этой границы было затруднено из-за сильной региональности большинства биостратиграфических маркеров и отсутствия каких-либо хемостратиграфических событий, таких как отклонения изотопов (большие внезапные изменения соотношений изотопов ), которые можно было бы использовать для определения или корреляции границы.. Calpionellids, загадочная группа планктонных протистов с урна-образный кальцитовых испытаний кратко обильны в течение последней юры до раннего мела, было предложено представить наиболее перспективных кандидатов для фиксации юры-мела границы В частности, первое появление Calpionella ALPINA, со - совпадает с основанием одноименной подзоны Альпина, что было предложено в качестве определения основания мелового периода. Рабочее определение границы часто рассматривалось как первое появление аммонита Strambergella jacobi, ранее относившегося к роду Berriasella, но его использование в качестве стратиграфического индикатора подвергалось сомнению, поскольку его первое появление не коррелирует с появлением C. альпина.
Киммериджа Глина и их эквиваленты являются основным источником роком для нефти в Северном море. Аравийский межшельфовый бассейн, образовавшийся в период средней и поздней юры, является местом расположения крупнейших в мире запасов нефти, включая месторождение Гавар, крупнейшее в мире месторождение нефти. Отложения юрского возраста Саргелу и Наокелекан являются основными нефтематеринскими породами Ирака. Более 1500 гигатонн запасов юрского угля находится на северо-западе Китая, в основном в бассейне Турфан-Хами и бассейне Ордос.
Основные ударные кратеры включают кратер Мороквенг, кратер диаметром 70 км, похороненный под пустыней Калахари на севере Южной Африки. Воздействие датируется границей юры и мела, около 145 млн лет назад. Было высказано предположение, что кратер Мороквенг сыграл роль в круговороте юрско-мелового перехода. Другой крупный ударный кратер - кратер Пучеж-Катунки диаметром 40–80 км, погребенный под Нижегородской областью, Россия. Удар был датирован синемурием, примерно 192–196 млн лет назад.
 Пангея в начале юрского периода
Пангея в начале юрского периода 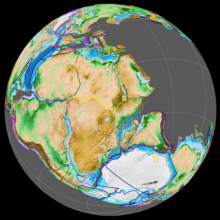 Распад Гондваны произошел во время поздней юры, в результате чего открылся Индийский океан.
Распад Гондваны произошел во время поздней юры, в результате чего открылся Индийский океан. В начале юрского периода все основные массивы суши в мире были объединены в суперконтинент Пангея, который во время ранней юры начал распадаться на северный суперконтинент Лавразию и южный суперконтинент Гондвану. Рифт между Северной Америкой и Африкой был первым, начавшимся в ранней юре и связанным с размещением Магматической провинции Центральной Атлантики.
В юрский период Северная Атлантика оставалась относительно узкой, тогда как Южная Атлантика не открывалась до мелового периода. Континенты были окружены Панталассой с океаном Тетис между Гондваной и Азией. В конце триаса в Европе произошла морская трансгрессия, затопившая большую часть центральной и западной Европы, превратив ее в архипелаг островов, окруженных мелководными морями. В юрский период и Северный, и Южный полюс были покрыты океанами. Начиная с ранней юры, Северный океан был связан с протоатлантическим океаном «коридором викингов» или трансконтинентальным Лавразийским морским путем, проходом между Балтийским щитом и Гренландией шириной несколько сотен километров.
Мадагаскар и Антарктида начали отделяться от Африки в конце ранней юры в связи с извержением крупных вулканических провинций Кару-Феррар, открывшим западную часть Индийского океана и начавшим фрагментацию Гондваны. В начале юрского периода Северная и Южная Америка оставались связанными, но к началу поздней юры они разделились, образуя Карибский морской путь, который соединил северную часть Атлантического океана с восточной частью Панталасса. Палеонтологические данные свидетельствуют о том, что морской путь был открыт с ранней юры.
В рамках Неваданской орогении, которая началась в триасовом периоде, Океан Кэш-Крик закрылся, и различные террейны, включая большой террейн Врангеллии, приросли к западной окраине Северной Америки. К средней юре Сибирская плита и Северо-Китайско-Амурский блок столкнулись, что привело к закрытию Монголо-Охотского океана.
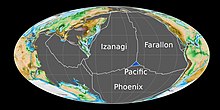 Формирование Тихоокеанской плиты в ранней юре.
Формирование Тихоокеанской плиты в ранней юре. Во время раннего юрского периода, около 190 миллионов лет назад, Тихоокеанская плита возникла в тройном стыке из Farallon, Феникс, и Идзанаги тектонических плит, из трех основных океанических плит из Панталассы. Ранее стабильный тройной стык превратился в нестабильную структуру, окруженную со всех сторон трансформными разломами из-за перегиба на одной из границ плит, что привело к образованию Тихоокеанской плиты в центре сочленения. В период от средней до ранней поздней юры Сандэнс Сивэй, мелководное эпиконтинентальное море, покрывало большую часть северо-запада Северной Америки.
 Grainstone с кальцитовыми ooids и шпатовым кальцитом цементом; Свита Кармель, средняя юра, южная часть штата Юта, США
Grainstone с кальцитовыми ooids и шпатовым кальцитом цементом; Свита Кармель, средняя юра, южная часть штата Юта, США По оценкам, эвстатический уровень моря был близок к нынешним уровням в течение геттангского и синемюрского периодов, поднялся на несколько десятков метров в течение позднего синемура – плинсбаха, прежде чем к позднему плинсбаху снизился до почти современных уровней. Похоже, что в тоарский период происходил постепенный подъем до пика ~ 75 м над нынешним уровнем моря. Во время последней части тоара уровень моря снова упал на несколько десятков метров. Он постепенно поднимался с ааленского яруса и далее, за исключением падений на несколько десятков метров в байосе и вокруг границы келловея и оксфорда, достигая, возможно, 140 метров над нынешним уровнем моря на границе кимериджа и титона. Уровень моря падает в конце титона, возможно, примерно до 100 метров, прежде чем подняться до примерно 110 метров на границе титона и берриаса.
Уровень моря в пределах долгосрочных трендов в юрском периоде был циклическим с 64 колебаниями, 15 из которых превышали 75 метров. Наиболее заметная цикличность в юрских породах относится к четвертому порядку с периодичностью примерно 410 000 лет.
В течение ранней юры мировые океаны перешли от арагонитового моря к химическому составу кальцитового моря, что способствовало растворению арагонита и осаждению кальцита. Подъем известнякового планктона во время средней юры глубоко изменил химию океана, при этом отложение биоминерализованного планктона на дно океана действует как буфер против крупных выбросов CO 2.
Климат юрского периода в целом был теплее, чем сейчас, примерно на 5-10 ° C, а содержание углекислого газа в атмосфере, вероятно, в четыре раза выше. Вероятно, леса росли возле полюсов, где было теплое лето и холодная, иногда снежная зима; маловероятно, что ледяные щиты были, учитывая высокие летние температуры, которые препятствовали накоплению снега, хотя могли быть и горные ледники. Камни и глендониты на северо-востоке Сибири в период от ранней до средней юры указывают на холодные зимы. Глубины океана были, вероятно, на 8 ° C теплее, чем сейчас, а коралловые рифы выросли на 10 ° широты дальше к северу и югу. Зона межтропической конвергенции, вероятно, существовала над океанами, в результате чего образовались большие площади пустынь и кустарников в более низких широтах между 40 ° северной широты и южной широты экватора. Биомы тропических лесов и тундры, вероятно, были редкими или отсутствовали.
Начало юры, вероятно, ознаменовалось термическим всплеском, соответствующим триасово-юрскому вымиранию и извержению Магматической провинции Центральной Атлантики. Первая часть юры была отмечена интервалом похолодания ранней юры между 199 и 183 миллионами лет назад. Было высказано предположение, что оледенение присутствовало в Северном полушарии во время плейнсбаха. В начале тоара произошел всплеск глобальных температур примерно на 4–8 ° C, соответствующий аноксическому событию в тоарском океане и извержению крупных вулканических провинций Кару-Феррар на юге Гондваны, с теплым интервалом, простирающимся до конец тоара около 174 миллионов лет назад.
Во время тоарского потепления температура поверхности океана, вероятно, превышала 30 ° C, а экваториальные и субтропические (30 ° с.ш. – 30 ° ю.ш.) регионы, вероятно, были чрезвычайно засушливыми, а температура внутри Пангеи, вероятно, превышала 40 °. С. За тоарским периодом потепления следует интервал похолодания в средней юре между 174 и 164 миллионами лет назад. Затем следует кимериджский теплый интервал между 164 и 150 миллионами лет назад. Внутри Пангеи сезонные колебания были менее серьезными, чем в предыдущие теплые периоды, поскольку расширение Центральной Атлантики и западной части Индийского океана предоставило новые источники влаги. Конец юры был отмечен интервалом похолодания титона – раннего баррема, начавшимся 150 миллионов лет назад и продолжавшимся до раннего мела.
Тоарское океаническое аноксическое событие (TOAE), также известное как событие Дженкинса, было эпизодом широко распространенной океанической аноксии в начале тоарской эры, c. 183 Mya. Он отмечен всемирно задокументированным выбросом отрицательных изотопов углерода с высокой амплитудой, а также отложением черных сланцев и исчезновением и коллапсом морских организмов, производящих карбонаты, что связано со значительным повышением глобальной температуры.
TOAE часто связывают с извержением крупных вулканических провинций Кару-Феррар и связанным с этим увеличением концентрации углекислого газа в атмосфере, а также с возможным сопутствующим выбросом клатратов метана. Это, вероятно, ускорило гидрологический цикл и увеличило силикатное выветривание. Затронутые группы включают аммонитов, остракод, фораминифер, брахиопод, двустворчатых моллюсков и книдарий, причем два последних отряда брахиопод, несущих шпильки, - Spiriferinida и Athyridida - вымирают. Хотя это событие оказало значительное влияние на морских беспозвоночных, на морских рептилий оно не повлияло. Во время TOAE бассейн Сычуань был преобразован в гигантское озеро, вероятно, в три раза больше современного озера Верхнее, представленное пачкой Дааньчжай из формации Цзилюцзин. Озера, вероятно, поглощенных ~460 гигатонн (GT) органического углерода и ~1,200 Гт неорганического углерода в ходе мероприятия. PH морской воды, который уже существенно снизился до события, немного увеличился на ранних стадиях TOAE, прежде чем упасть до самой низкой точки примерно к середине события. Это закисление океана, вероятно, вызвало коллапс производства карбонатов.
Переход к концу юрского периода первоначально считался одним из восьми массовых вымираний, но теперь считается сложным периодом круговорота фауны с увеличением разнообразия одних групп и сокращением других, хотя свидетельства этого в основном европейские, вероятно контролируется изменениями эвстатического уровня моря.
Нет свидетельств массового вымирания растений на рубеже триаса и юры. На границе триаса и юры в Гренландии данные о спороморфах (пыльца и споры) предполагают полный круговорот цветков. Анализ цветочных сообществ макрофоссилий в Европе показывает, что изменения в основном были вызваны местной экологической сукцессией. В конце триаса Peltaspermaceae вымерли в большинстве частей мира, а Lepidopteris сохранились до ранней юры в Патагонии. Dicroidium, семенной папоротник, который был доминирующей частью цветочных сообществ Гондваны в течение триаса, также пришел в упадок на границе триаса и юры, сохранившись в качестве реликта в Антарктиде до синемурийского периода.
 Окаменелая шишка Araucaria mirabilis из средней юры Аргентины
Окаменелая шишка Araucaria mirabilis из средней юры Аргентины Хвойные деревья составляли доминирующий компонент юрских флор. Поздний триас и ранняя юра были основным периодом диверсификации хвойных пород, при этом большинство современных групп хвойных пород появилось в летописи окаменелостей к концу юрского периода, развившись от вольциаловых предков.
Первые недвусмысленные упоминания о хвойных деревьях араукария относятся к ранней юре, а те, которые тесно связаны с современным родом Araucaria, были широко распространены в обоих полушариях к средней юре.
В юрском периоде также широко распространено вымершее семейство Cheirolepidiaceae, которое часто распознается по их очень характерной пыльце Classopolis. Представители юрского периода включают пыльцевую шишку Classostrobus и семенную шишку Pararaucaria. Хвойные деревья араукариан и хейролепидиевые часто встречаются вместе.
Самая старая окончательная запись семейства кипарисовых ( Cupressaceae ) - это Austrohamia minuta из ранней юры (плиенсбахи) Патагонии, известная по многим частям растения. Репродуктивные структуры Austrohamia имеют сильное сходство с таковыми у примитивных живых кипарисов родов Taiwania и Cunninghamia. К средней и поздней юре Cupressaceae были многочисленны в теплых умеренно-тропических регионах Северного полушария, наиболее широко представленные родом Elatides.
Представители вымершего рода Schizolepidopsis, которые, вероятно, представляют стеблевую группу семейства сосновых ( Pinaceae ), были широко распространены в Евразии в юрский период. Самая старая однозначная находка сосновых - это сосновая шишка Eathiestrobus, известная из поздней юры (кимеридж) в Шотландии, которая остается единственной известной однозначной окаменелостью группы до мелового периода. Несмотря на то, что Eathiestrobus является самым ранним известным представителем Pinaceae, он, по-видимому, является членом пиноидной клады семейства, что позволяет предположить, что первоначальное разнообразие Pinaceae произошло раньше, чем это было обнаружено в летописи окаменелостей.
В течение ранней юры во флоре средних широт Восточной Азии преобладали вымершие лиственные широколиственные хвойные растения Podozamites, которые, по-видимому, не были тесно связаны ни с одним из ныне живущих семейств хвойных. Его ареал простирался на север до полярных широт Сибири, а затем сужался к северу в средней и поздней юре, что соответствовало увеличению засушливости региона.
Самая ранняя находка семейства тисовых ( Taxaceae ) - Palaeotaxus rediviva из геттангского яруса в Швеции, предположительно тесно связана с живущим Austrotaxus, в то время как Marskea jurassica из средней юры Йоркшира, Англия, и материалы из келловейско-оксфордского пласта Даохугоу. в Китае, как полагают, тесно связаны с Amentotaxus, причем последний материал отнесен к современному роду, что указывает на то, что Taxaceae существенно расширились к концу юрского периода.
Podocarpaceae, в настоящее время в основном приуроченные к Южному полушарию, встречались в Северном полушарии во время юрского периода. Примеры включают Podocarpophyllum из ранней и средней юры в Центральной Азии и Сибири, Scarburgia из средней юры Йоркшира и Harrisiocarpus из юрского периода в Польше.
 Листья Ginkgo huttonii из средней юры Англии
Листья Ginkgo huttonii из средней юры Англии Ginkgoales, единственным живым видом которых является Ginkgo biloba, были более разнообразны в юрский период: они были одними из наиболее важных компонентов юрских флор Евразии и были адаптированы к широкому спектру климатических условий. Самые ранние представители рода Ginkgo, представленные овуляционными и пыльцевыми органами, сходными с таковыми у современных видов, известны из средней юры в Северном полушарии. Несколько других линий гинкгоалий известны из юрских пород, в том числе Yimaia, Grenana, Nagrenia и Karkenia. Эти линии связаны с гинкго- подобными листьями, но отличаются от живых и ископаемых представителей гинкго наличием по-разному устроенных репродуктивных структур. Умальтолепис, исторически считающийся гинкгоалеем, и владимария из юрского периода Азии имеют ленточноподобные листья гинкго ( Pseudotorrelia ) с очень отчетливыми репродуктивными структурами, сходными с таковыми у папоротников семян пельтасперма и користосперма; они размещены в отдельном порядке « Владимариалес», что может быть связано с гинкгоалесами.
Bennettitales - это группа семенных растений, широко распространенных в мезозое, которые достигли пика разнообразия в триасовый и юрский периоды. Листва Bennettitales очень похожа на листву саговников до такой степени, что их невозможно надежно отличить только на основе морфологии. Листья Bennettitales можно отличить от листьев саговников по разному расположению устьиц, и эти две группы не считаются тесно связанными. Все юрские Bennettitales принадлежат к группе Williamsoniaceae, которые росли как кустарники и небольшие деревья. Полагают, что у Williamsoniaceae была разветвленная форма ветвления, аналогичная привычке живых Banksia, и они были приспособлены к выращиванию в открытых местообитаниях с плохими условиями содержания питательных веществ в почве. Bennettitales демонстрируют сложные репродуктивные структуры, похожие на цветы, которые, как считается, были опылены насекомыми. Предполагается, что несколько групп насекомых с длинным хоботком, включая вымершие семейства, такие как златоглазки каллиграмматид, и современные насекомые, такие как акроцеридные мухи, были опылителями беннеттитовых, питаясь нектаром, произведенным шишками беннеттитали.
Цикады достигли пика своего разнообразия в юрский и меловой периоды. Несмотря на то, что мезозой иногда называют «веком саговников», считается, что саговники были относительно второстепенным компонентом средне-мезозойских флор, их листву путали с листвой Bennettitales и других «цикадофитов». В Nilssoniales есть листья морфологически сходные с цикады, и часто считаются цикады или Cycad родственники, но было установлено, что различны по химическим признакам, и, возможно, более тесно связаны с беннеттитовых.Считается, что саговники в основном обитали в тропических и субтропических широтах на протяжении всей своей эволюционной истории. Отношения большинства мезозойских саговников с живыми группами неоднозначны. Семена юрского периода в Англии и Хайда-Гвайи, Канада, являются ранними представителями Cycadaceae, единственным живым родом которых является Cycas. Семена, обнаруженные в кишечнике динозавра Isaberrysaura из средней юры Аргентины, относятся к Zamiineae, в которую входят все другие живущие роды саговников. Современные саговники опыляются жуками, и считается, что такая ассоциация сформировалась к ранней юре.
Несмотря на то, что было несколько заявленных записей, нет общепринятых отчетов юрского периода о цветковых растениях, которые составляют 90% живых видов растений, а свидетельства окаменелостей предполагают, что в течение следующего мелового периода группа диверсифицировалась.
Gnetophyte Protognetum из средней юры Китая демонстрирует характеристики как гнетума и эфедр, и помещается в монотипической семье Protognetaceae.
 Sagenopteris phillipsi ( Caytoniales ) из средней юры Йоркшира, Англия
Sagenopteris phillipsi ( Caytoniales ) из средней юры Йоркшира, Англия «Семенные папоротники» ( Pteridospermatophyta ) - собирательный термин для обозначения разрозненных линий папоротниковых растений, которые дают семена, но имеют неопределенное родство с живыми группами семенных растений. Выдающаяся группа юрских семенных папоротников - это Caytoniales, которые достигли своего зенита в юрский период и широко известны в Северном полушарии, хотя записи в Южном полушарии остаются редкими. Из - за их ягодное -like семенных капсул, они часто были предложены, были тесно связаны или, возможно, предок цветущих растений, но доказательства этого неубедительно. Записи о користоспермах, группе семенных папоротников, прежде всего известной из триаса Гондваны, становятся редкими после триаса и упадка дикроидиума, хотя, похоже, есть юрские записи группы из Северного полушария.
Czekanowskiales, также известные как Leptostrobales, представляют собой группу семенных растений неопределенного родства с устойчивыми сильно рассеченными листьями на лиственных коротких побегах, покрытых чешуевидными листьями, известными от позднего триаса (возможно, поздней перми) до мелового периода. Считается, что они имели вид деревьев или кустарников и составляли заметный компонент мезозойских умеренных и теплых флор Северного полушария. Род Phoenicopsis был широко распространен во флорах ранне-средней юры Восточной Азии и Сибири.
Пентоксиловой, небольшая, но явно отличается группа семенных растений неясного сродства, впервые появилась в юрском период. Их распространение, по-видимому, ограничивалось Гондваной.
Живые семейства папоротников, широко распространенные в юрский период, включают Dipteridaceae, Matoniaceae, Gleicheniaceae, Osmundaceae и Marattiaceae. Многоножковые, которые составляют 80% от живого папоротника разнообразия, не имеют никаких записей из юры и, как полагает, диверсифицируют в меле, хотя широкого Jurassic травянистого папоротник рода Coniopteris, исторически интерпретирован как близкий родственник древовидных папоротников семьи Dicksoniaceae, недавно был переосмыслен как ранний родственник группы.
Древовидные папоротники, группа, содержащая самый современные древовидные папоротники, появившиеся в позднем юры, в лице представителей рода Cyathocaulis, которые предлагают быть ранними членами Cyatheaceae на основе кладистического анализа. Существует лишь несколько возможных записей о Hymenophyllaceae из юрского периода, включая Hymenophyllites macrosporangiatus из юрского периода в России.
Самые старые останки современных хвощей рода Equisetum впервые появляются в ранней юре, представленных Equisetum dimorphum из ранней юры Патагонии и Equisetum laterale из ранней и средней юры Австралии. Силикатные остатки Equisetum thermale из поздней юры Аргентины демонстрируют все морфологические признаки современных представителей этого рода. Предполагаемый раскол между Equisetum bogotense и всеми остальными живыми Equisetum произошел не позднее ранней юры.
Полынь, практически идентичная современным видам, известна с юрского периода. Isoetites rolandii из средней юры Орегона - самый ранний известный вид, представляющий все основные морфологические особенности современных Isoetes. Более примитивные формы, такие как наторстиана, сохраняющие удлиненный стебель, сохранились до раннего мелового периода.
Считается, что мох Kulindobryum из средней юры России, связанный с костями динозавров, является родственником Splachnaceae, которые растут на каракасах животных. Считается, что Bryokhutuliinia из того же региона связана с Dicranales. Считается, что Heinrichsiella из юрского периода Патагонии принадлежит либо к Polytrichaceae, либо к Timmiellaceae.
Печеночник Pellites hamiensis из средней юры формирования Xishanyao Китая является самой старой записью семьи Pelliaceae. Считается, что Pallaviciniites sandaolingensis из того же месторождения относится к подклассу Pallaviciniineae в пределах Pallaviciniales. Ricciopsis sandaolingensis, также из того же месторождения, является единственным юрским летописцем Ricciaceae.
 Голотип образец Platysuchus, telosaurid талаттозухии
Голотип образец Platysuchus, telosaurid талаттозухии Триасово-юрское вымирание уничтожило разнообразие псевдозухий, при этом крокодиломорфы, возникшие в раннем позднем триасе, были единственной группой псевдозухий, которые выжили, а все остальные, включая травоядных этозавров и хищных « рауизухий », вымерли. Морфологическое разнообразие крокодиломорфов во время ранней юры было примерно таким же, как у позднетриасовых псевдозухий, но они занимали разные области морфопространства, что свидетельствует о том, что они занимали разные экологические ниши по сравнению со своими триасовыми собратьями и что крокодиломорфы подвергались обширной и быстрой радиации. в течение этого интервала. В то время как живые крокодилы приурочены к образу жизни хищников из засад в водной среде, юрские крокодиломорфы демонстрировали широкий спектр жизненных привычек. Безымянный протозухид, известный по зубам из ранней юры Аризоны, представляет собой самого раннего известного травоядного крокодиломорфа, адаптацию, которая появлялась несколько раз в течение мезозоя.
Талаттозухии, клады преимущественно морских крокодиломорфов, впервые появились в раннем юры и стала заметной частью морских экосистем. Внутри Thalattosuchia Metriorhynchidae стали хорошо приспособлены к жизни в открытом океане, включая превращение конечностей в ласты, развитие хвостовой двуустки и гладкую безчешуйную кожу. Морфологическое разнообразие крокодиломорфов в раннюю и среднюю юрское время было относительно низким по сравнению с таковым в более поздние периоды времени, и преобладали наземные мелкотелые, длинноногие сфенозухи, ранние крокодилиформные и талаттозухи. Neosuchia, основная группа крокодиломорфов, впервые появилась в начале-средней юры. Neosuchia представляет собой переход от наземного образа жизни предков к пресноводной водной экологии, аналогичной экологии современных крокодилов. Время происхождения Neosuchia оспаривается. Самая старая запись Neosuchians было предложено, чтобы быть Calsoyasuchus, от ранней юры Аризоны, который во многих анализах, извлеченного в качестве самого раннего разветвлений члена семьи neosuchian Goniopholididae, которые коренным образом изменяет времена диверсификации для крокодиломорфы. Тем не менее, это размещение оспаривается, и некоторые анализы обнаружили его за пределами Неозухии, что позволило бы разместить самые старые записи о Неозухии в средней юре. Было высказано предположение, что разанандронгоб из средней юры Мадагаскара представляет собой старейшее упоминание о нотосучиях, преимущественно гондванской кладе, состоящей в основном из наземных крокодиломорфов, известных еще с мелового и кайнозойского периодов.
 Thalassemys, thalassochelydian морских черепах известный из позднего юрского Германии
Thalassemys, thalassochelydian морских черепах известный из позднего юрского Германии Стеблевая группа черепах ( Testudinata ) разнообразилась в юрский период. Юрские стволовые черепахи принадлежат к двум прогрессивно более развитым кладам, Mesochelydia и Perichelydia. Считается, что родовое состояние мезохелидийцев - водное, в отличие от наземных для семенников. Две современные группы черепах ( Testudines ), Pleurodira и Cryptodira, разошлись в средней юре. Самые старые известные плевродиры, Platychelyidae, известны с поздней юры. Thalassochelydia, разнообразное происхождение морских черепах, как известно, от позднего юрского Европы и Южной Америки.
Ринхоцефалы (единственный живой представитель - туатара ) получили глобальное распространение к началу юрского периода. Rhynchocephalians достигли наивысшего морфологическое разнообразие в их эволюционной истории в юрский период, занимающий широкий диапазон образа жизни, в том числе водные pleurosaurs с длинной змеи, как тела и уменьшенных конечностей, специализированные растительноядные eilenodontines, а также Oenosaurus, которые имели широкие зубные пластины указывает на дурофагию. Считается, что циносфенодон из ранней юры Мексики является одним из ближайших известных родственников ныне живущих туатар. Ринхоцефалы исчезли из Азии после ранней юры. Последний общий предок современных чешуекрылых (включая ящериц и змей ), по оценкам, жил около 190 миллионов лет назад во время ранней юры, при этом основные расхождения между чешуйчатыми линиями, по оценкам, произошли в период ранней и средней юры. Скваматы впервые появляются в летописи окаменелостей в средней юре и включали ранних представителей змеиной линии ( Ophidia ) и Scincomorpha, хотя многие чешуйки юрского периода имеют неясные отношения с живыми группами. Eichstaettisaurus из поздней юры Германии считается одним из первых родственников гекконов и демонстрирует приспособления для лазания. Дорсетизавр из поздней юры Северной Америки и Европы представляет собой старейшее широко признанное упоминание об ангиморфах. Марморетта из средней юры Британии представляет собой выжившего недавно лепидозавроморфа за пределами как Rhynchocephalia, так и Squamata.

Vadasaurus herzogi, рынхоцефал из верхнеюрских известняков Зольнхофена в Германии.

Homeosaurus maximiliani, рынхоцефал из Зольнхофенского известняка

Pleurosaurus,, водное rhynchocephalian от позднего юрского Европы

Eichstaettisaurus schroederi, вымершая ящерица из Зольнхофенского известняка.
Самые ранние известные останки Choristodera, группы пресноводных водных рептилий с неопределенным родством с другими группами рептилий, найдены в средней юре. Из юры известны всего два рода хористодеров. Один из них - это маленькие ящерицы Cteniogenys, которые считаются наиболее базальными из известных хористодеров; он известен от средней до поздней юры Европы и поздней юры Северной Америки; аналогичные остатки известны также из верхней средней юры Кыргызстана и Западной Сибири. Другой - Coeruleodraco из поздней юры Китая, который представляет собой более продвинутый хористодер, хотя все еще маленький и похожий на ящерицу по морфологии.
 Ископаемое Ichthyosaurus communis в Музее естественной истории в Лондоне
Ископаемое Ichthyosaurus communis в Музее естественной истории в Лондоне Ихтиозавры пережили узкое место в эволюции во время вымирания в конце триаса, когда вымерли все, не относящиеся к неоихтиозаврам. Ихтиозавры достигли пика видового разнообразия во время ранней юры, с множеством морфологий, включая огромного хищника-хищника Темнодонтозавра и меч- рыбоподобного Eurhinosaurus, хотя раннеюрские ихтиозавры были значительно менее разнообразны морфологически, чем их триасовые собратья. На границе раннего и среднего триаса, между концом тоара и началом байоса, большинство линий ихтиозавров, по-видимому, вымерли, с первым появлением Ophthalmosauridae, клада, которая с тех пор охватывала почти всех ихтиозавров., в раннем байосе. Офтальмозавриды были разнообразны к поздней юре, но не смогли заполнить многие ниши, которые были заняты ихтиозаврами в раннеюрский период.
 Rhomaleosaurus cramptoni в Музее естественной истории
Rhomaleosaurus cramptoni в Музее естественной истории Плезиозавры возникли в конце триаса (рета). К концу триаса все другие зауроптериги, включая плакодонтов и нотозавров, вымерли. По крайней мере шесть линий плезиозавров пересекли границу триаса и юры. Плезиозавры были разнообразны уже в самом раннем юрском периоде, при этом большинство плезиозавров в голубых лиасах геттангийского возраста принадлежали к ромалеозавридам. Ранние плезиозавры, как правило, были небольшого размера, а размер тела увеличивался до тоара. Похоже, что на границе ранней и средней юры произошел сильный круговорот с исчезновением ранее широко распространенных ромалеозаврид и микроклеидид после конца тоара и первым появлением доминирующей клады плезиозавров второй половины юры. Cryptoclididae, во байосе. В средней юре произошла эволюция талассофонных плиозавров с короткой шеей и крупноголовыми талассофоническими плиозаврами от предков с короткой шеей и длинной шеей. У некоторых талассофоновых плиозавров, таких как некоторые виды Pliosaurus, были черепа до двух метров в длину, а длина тела оценивалась примерно в 10–12 метров, что делало их высшими хищниками позднеюрских океанов. Плезиозавры вторглись в пресноводную среду в юрский период с неопределенными останками мелких плейзозавров, известных из пресноводных отложений юрского периода в Китае и Австралии.
 Скелет Rhamphorhynchus muensteri в музее Тейлера, Харлем
Скелет Rhamphorhynchus muensteri в музее Тейлера, Харлем Впервые птерозавры появились в позднем триасе. Основное излучение юрских птерозавров - это Rhamphorhynchidae, которые впервые появились в конце ранней юры (тоар); они считаются рыбоядными. Анурогнатиды, впервые появившиеся в средней юре, обладали короткими головами и густо покрытыми мехом телами и, как полагают, были насекомоядными. Производные монофенестратановые птерозавры, такие как вуконгоптериды, появились в конце средней юры. Развитые короткохвостые птеродактилоиды впервые появились на границе средней и поздней юры. К юрским птеродактилоидам относятся ктенохазматиды, такие как Ctenochasma, с близко расположенными игольчатыми зубами, которые предположительно использовались для питания через фильтр. У причудливого позднеюрского ктенохазматоида Cycnorhamphus была челюсть с зубами только на кончиках, с изогнутыми челюстями, как у живых аистов открытого клюва, которые, возможно, использовались для удержания и раздавливания твердых беспозвоночных.
Динозавры, которые морфологически разнообразились в позднем триасе, испытали значительное увеличение разнообразия и численности в ранней юре после вымирания в конце триаса и исчезновения других групп рептилий, став доминирующими позвоночными в наземных экосистемах. Чилезавр, морфологически аберрантный травоядный динозавр из поздней юры Южной Америки, имеет неопределенные отношения к трем основным группам динозавров, которые были обнаружены как член всех трех в различных анализах.
Развитые тероподы, принадлежащие к Neotheropoda, впервые появились в позднем триасе. Базальные неотроподы, такие как целофизоиды и дилофозавры, существовали до ранней юры, но вымерли к средней юре. Самые ранние авеространы появляются в раннем юрском периоде, причем самым ранним известным представителем Ceratosauria является Saltriovenator из раннего синемурийского периода (199,3–197,5 миллионов лет назад) Италии. Необычный цератозавр Limusaurus из поздней юры Китая питался травоядными животными, а взрослые особи имели беззубые клювые челюсти, что сделало его самым ранним известным тероподом, перешедшим из хищной диеты предков. Самые ранние представители Tetanurae появились в конце ранней юры или ранней средней юре. Megalosauridae представляет старейшее излучение тетануров, первое появление в Европе в байосе. Предполагается, что старейшим представителем Allosauroida является Asfaltovenator из средней юры Южной Америки. Целурозавры впервые появились в среднем юрском периоде, включая ранних тираннозавров, таких как Процератозавр из Бата в Британии. Некоторые целурозавры из поздней юры Китая, включая Shishugounykus и Haplocheirus предлагаются представлять ранние alvarezsaurs, однако, это было поставлена под сомнение. Скансориоптеригиды, группа маленьких пернатых целурозавров с перепончатыми крыльями, напоминающими крылья летучей мыши для планирования, известны от средней до поздней юры Китая. Предполагается, что самая старая находка троодонтид - это Hesperornithoides из поздней юры Северной Америки. Остатки зубов, предположительно принадлежащие дромеозаврам, известны с юрского периода, но до мелового периода не известны останки останков.

Скелет Ceratosaurus, ceratosaurid от позднего юрского периода Северной Америки

Скелет монолофозавра, базального тетанурана из средней юры Китая

Восстановление И Ци, скансориоптеригида от средней до поздней юры Китая
 Archeopteryx lithographica из поздней юры (титона) Германии
Archeopteryx lithographica из поздней юры (титона) Германии Самые ранние птичьи птицы, в том числе птицы и их предки, появляются в период от средней до поздней юры и окончательно представлены археоптериксом из поздней юры Германии. Avialans принадлежит к кладе Paraves в составе целурозавров, в которую также входят дромеозавры и троодонтиды. Anchiornithidae от средне-поздней юры Евразии часто предлагает быть avialans, но также в качестве альтернативы найдены в виде отдельной родословной паравесов.
 Скелет гетеродонтозавра, примитивного орнитиша из ранней юры Южной Африки
Скелет гетеродонтозавра, примитивного орнитиша из ранней юры Южной Африки Самые ранние окончательные орнитисхий появляются во время раннего юрского периода, представленная базальными орнитисхий как Lesothosaurus, heterodontosaurids и первых членов тиреофоры. Самые ранние представители анкилозаврии и стегозаврии появляются в среднем юрском периоде. Базальный neornithischian Kulindadromeus из средней юры России указывает на то, что по крайней мере некоторые орнитисхи были покрыты проточеками. Самые ранние представители анкилополлексии, которые стали заметными в меловом периоде, появились во время поздней юры и были представлены двуногими формами, такими как камптозавр. Цератопсы впервые появились в поздней юре Китая, представленные представителями Chaoyangsauridae.
 Скелет Mamenchisaurus sinocanadorum из средней-поздней юры Китая
Скелет Mamenchisaurus sinocanadorum из средней-поздней юры Китая Зауроподы стали доминирующими крупными травоядными в наземных экосистемах в юрский период. Некоторые юрские зауроподы достигли гигантских размеров, став самыми крупными организмами, когда-либо жившими на суше.
Базальные двуногие зауроподоморфы, такие как массоспондилиды, продолжали существовать до ранней юры, но вымерли к началу средней юры. Четвероногие зауропоморфы появились в позднем триасе. Четвероногий Ledumahadi с самим ранним юрским Южной Африкой достиг оценочного вес 12 тонн, намного выше других известных завроподоморфов базальных. Гравизаврские зауроподы впервые появились в раннем юрском периоде, причем самой старой окончательной находкой был вулканодон из Зимбабве, вероятно, синемурийского возраста. Евзауроподы впервые появились в конце ранней юры (тоар) и разнообразились в течение средней юры; к ним относятся цетиозавриды, туриазавры и маменчизавры. Самый ранний известный представитель Neosauropoda - Lingwulong, диплодокоид дикреозаврид из ранней и средней юры Китая. Неозауроподы расселились по всему миру в поздней юре. В средней-поздней юре впервые появились производные группы неозауроподов, включая Brachiosauridae и Diplodocidae.
 Скелет Karaurus sharovi, саламандры стебельной группы от средней до поздней юры Казахстана
Скелет Karaurus sharovi, саламандры стебельной группы от средней до поздней юры Казахстана Разнообразие темноспондилей постепенно сокращалось в течение позднего триаса, и только брахиопоиды выжили до юры и далее. Члены семьи Brachyopidae известны из юрских отложений в Азии, в то время как chigutisaurid сидеропс известно из ранней юры Австралии. Разнообразие современных лиссамфибий началось в юрский период. Раннеюрский просалирус считался первым родственником лягушек с морфологией, способной прыгать, как живые лягушки. Морфологически узнаваемые стволовые лягушки, такие как южноамериканский нотобатрахус, известны со средней юры. В то время как самые ранние земноводные саламандры известны из триаса, саламандры коронной группы впервые появляются в Евразии в период от средней до поздней юры вместе с родственниками стволовой группы. Считается, что многие юрские саламандры стволовой группы, такие как Marmorerpeton и Kokartus, были неотениками. Ранние представители саламандр коронной группы включают Chunerpeton, Pangerpeton и Linglongtriton от средней до поздней юрской биоты Янляо Китая. Они принадлежат к Cryptobranchoidea, который содержит живых азиатских и гигантских саламандр. Бейянерпетон и Цинлунтритон из одной и той же биоты считаются первыми членами Саламандройда, группы, в которую входят все остальные живые саламандры. Саламандры расселились в Северной Америке к концу юрского периода, о чем свидетельствует иридотритон, обнаруженный в позднеюрской формации Моррисон. Самый старый бесспорный стебель- цецилий - это раннеюрская эокаецилия из Аризоны. Четвертая группа лиссамфибий, вымершие albanerpetontids, впервые появились в средней юре и представлены Anoualerpeton priscus из бата в Британии, а также неопределенными останками из отложений того же возраста во Франции и формации Anoual в Марокко.
 Henkelotherium, вероятный древесный диолестоид из поздней юры Португалии
Henkelotherium, вероятный древесный диолестоид из поздней юры Португалии Млекопитающие, произошедшие от цинодонтов в конце триаса, широко разнообразились в юрском периоде. Важные группы юрских млекопитающих включают Morganucodonta, Docodonta, Eutriconodonta, Dryolestida, Haramiyida и Multituberculata. В то время как большинство юрских млекопитающих известны исключительно по изолированным зубам и фрагментам челюстей, исключительно сохранившиеся останки свидетельствуют о разнообразии образа жизни. Докодонт Castorocauda был адаптирован к водной жизни, так же как утконос и выдры. У некоторых представителей Haramiyida и эвтриконодонтского племени Volaticotherini был патагиум, похожий на таковой у белок-летяги, позволяющий им парить в воздухе. Муравьеда -как млекопитающее фрутафоссор, неопределенной систематика, скорее всего, был специалист по колониальным насекомыми, подобно живым муравьедам. Australosphenida, группа, в которую входят современные монотремы, впервые появилась в средней юре Гондваны. Therian млекопитающие, представленные сегодня живыми плацентарных и сумчатых, появляются в начале поздней юры, представлен Juramaia, в плацентарные млекопитающих ближе к родословной плацентарных, чем сумчатых. Джурамая намного более развита, чем ожидалось для своего возраста, поскольку другие терианские млекопитающие не были известны до раннего мелового периода. Две группы цинодонтов, не относящихся к млекопитающим, сохранились и после конца триаса. Насекомоядные Tritheledontidae имеют несколько записей из ранней юры. Тритилодонтовые, herbiviorous группа cynodonts, которые впервые появились во рэте, имеет обильные записи из юры, в подавляющем большинстве из северного полушария.
Последний известный вид конодонтов, класс бесчелюстных рыб, чьи твердые зубчатые элементы являются ключевыми ископаемыми останками, окончательно вымер в ранней юре после более чем 300 миллионов лет эволюционной истории, причем асинхронное вымирание произошло сначала в Тетисе и на востоке страны. Панталасса и выжившие, сохранившиеся до самого раннего геттанга Венгрии и центральной Панталассы. Конодонты конца триаса были представлены лишь несколькими видами и постепенно сокращались в среднем и позднем триасе.
 Целакант из Зольнхофенского известняка
Целакант из Зольнхофенского известняка Двоякодышащие (Dipnoi) обитали в пресноводных средах обоих полушарий в юрский период. К родам относятся Ceratodus и Ptychoceratodus, которые более тесно связаны с живыми южноамериканскими и африканскими двоякодышащими рыбами, чем с двоякодышащими двояковыми рыбами из Квинсленда, и Ferganoceratodus из юрского периода в Азии, которые не имеют близкого родства ни с одной из групп ныне живущих двоякодышащих рыб. Моусонииды, морская и пресноводная / солоноватая группа целакантов, впервые появившаяся в Северной Америке в триасовый период, распространилась в Европу и Южную Америку к концу юрского периода. Морские Latimeriidae, которые содержат живые латимерии рода Latimeria, также присутствовали в юрском периоде и возникли в триасе.
 Голова и передний плавник Pachycormus, вымершей толстокожей рыбы
Голова и передний плавник Pachycormus, вымершей толстокожей рыбы Костные рыбы ( Actinopterygii ) были основными компонентами юрских пресноводных и морских экосистем. Амииформная рыба (которая сегодня включает только боуфин ) впервые появилась в раннем юрском периоде и была представлена Катуром из плинсбахского периода в Британии; после своего появления в западной части Тетиса, к концу юрского периода они распространились на Африку, Северную Америку, Юго-Восточную и Восточную Азию. Pycnodontiformes, которые впервые появились в западной части Тетиса в позднем триасе, распространились на Южную Америку и Юго-Восточную Азию к концу юрского периода, имея большое разнообразие в Европе в течение поздней юры. Костистые кости, составляющие более 99% современных актиноптеригий, впервые появились в триасовом периоде в западной части Тетиса; они претерпели серьезную диверсификацию, начавшуюся в поздней юре, с появлением в это время ранних представителей современных клад костистых, таких как Elopomorpha и Osteoglossoidei. В Pachycormiformes, группа рыб тесно связан с костистых, впервые появился в ранней юры и включала в себя как тунец -like хищными и фильтр кормления форм. Последний включал в себя самую крупную из когда-либо существовавших костных рыб: Лидсихтис, с предполагаемой максимальной длиной более 15 метров, известный от конца среднего до позднего юрского периода.
 Ископаемые из Palaeocarcharias, самая старая известная lamniform акула
Ископаемые из Palaeocarcharias, самая старая известная lamniform акула В течение ранней юры гибодонты, такие как Hybodus, были обычны как в морских, так и в пресноводных условиях; однако к поздней юре гибодонты стали второстепенными компонентами большинства морских сообществ, будучи в значительной степени вытесненными современными неозелахами, но оставались обычными в пресноводных и ограниченных морских средах. Neoselachii, в котором обитают все живые акулы и скаты, излучались начиная с ранней юры. Самые старые из известных Hexanchiformes происходят из ранней юры (плиенсбахи) Европы. Самый старый известный скат ( Batoidea ) - это Antiquaobatis из плинсбахского периода в Германии. Юрские батоиды, известные из полных останков, сохраняют консервативную морфологию, напоминающую гитарфиш. Самые старые известные родственники бычьей акулы ( Heterodontus) в отряде Heterodontiformes впервые появились в ранней юре, а представители живого рода появились в поздней юре. Акулы ковров (Orectolobiformes) впервые появились во тоаре, представленной Folipistrix и Annea из Европы. Самые старые известные скумбрийские акулы (Lamniformes) происходят из средней юры, представленные родом Palaeocarcharias, который имеет тело, похожее на прямолинейное тело, но имеет ключевые сходства в гистологии зубов с Lamniformes, включая отсутствие ортодентина. Самая старая находка акул-ангелов (Squatiniformes) - это Pseudorhina из оксфордско-титонского периода Европы. Самые старые известные останки Carcharhiniformes, крупнейшего отряда ныне живущих акул, впервые появляются в конце средней юры (бат) западной части Тетиса (Англия и Марокко). Известные зубные и исключительно сохранившиеся остатки трупов юрских кархаринообразных похожи на останки современных кошачьих акул. Synechodontiformes, вымершая группа акул, тесно связанных с Neoselachii, также была широко распространена в юрский период. Самые старые останки химер относятся к ранней юре Европы, а представители современного семейства Callorhinchidae появились в средней юре. В отличие от живых химер, они были найдены на мелководье. Тесно связаны Squaloraja и myriacanthoids также известны с юры Европы.
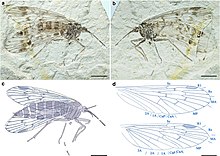 Lichnomesopsyche daohugouensis, вымерший мезопсихидный скорпион из поздней юры Китая
Lichnomesopsyche daohugouensis, вымерший мезопсихидный скорпион из поздней юры Китая По-видимому, на границе триаса и юры не было значительного исчезновения насекомых. Многие важные местонахождения окаменелостей насекомых известны из юрского периода Евразии, наиболее важными из которых являются формация Карабастау в Казахстане и различные месторождения биоты Янляо во Внутренней Монголии, Китай, такие как пласт Даохугоу, датируемые келловейем – оксфордом. Разнообразие насекомых стагнировало на протяжении всей ранней и средней юры, но во второй трети юрского периода темпы возникновения значительно увеличились, в то время как темпы исчезновения оставались неизменными. Возрастающее разнообразие насекомых в средней – поздней юре соответствует значительному увеличению разнообразия ротового аппарата насекомых. Средняя и поздняя юра была временем большого разнообразия жуков. Долгоносики впервые появляются в летописи окаменелостей в период от средней до поздней юры, но предполагается, что они возникли в период от позднего триаса до ранней юры. Самые старые из известных чешуекрылых (группа бабочек и мотыльков) известны на границе триаса и юры, с чешуей крыльев, принадлежащих подотряду Glossata, и бабочек класса Micropterigidae из отложений этого возраста в Германии. Современные представители как стрекоз, так и стрекоз также впервые появились в юрский период. Хотя современные представители не известны до кайнозоя, эктопаразитические насекомые, которые, как считается, представляют примитивных блох, принадлежащих к семейству Pseudopulicidae, известны из средней юры Азии. Эти насекомые существенно отличаются от современных блох, не имея специальной морфологии последних и более крупные. Самая ранняя группа Phasmatodea (палочников) - крылатые Susumanioidea, внешняя группа по сравнению с современными Phasmatodea - впервые появилась в средней юре. В это время также появился старейший представитель Mantophasmatidae (гладиаторов).
 Монголарахне из поздней юры Китая
Монголарахне из поздней юры Китая Лишь немногие из записей клещей известны с юры, в том числе Jureremus, в панцирных клещей, принадлежащих к семейству Cymbaeremaeidae известно из позднего юрского Великобритании и России, а член все еще живет orbatid род Hydrozetes из ранней юры Швеции. Появление пауков появилось в юрском периоде. Раннеюрское koponeni Сеппо может представлять собой стволовую группу Palpimanoidea. Eoplectreurys из средней юры Китая считается стволовой линией Synspermiata. Самый старый член семьи Archaeidae, Patarchaea, известен из средней юры Китая. Mongolarachne из средней юры Китая - один из крупнейших известных ископаемых пауков с ногами более 5 сантиметров. Единственный известный юрский скорпион - это Liassoscorpionides из ранней юры Германии, местонахождение неизвестно. Сборщики урожая Eupnoi ( Opiliones ) известны из средней юры Китая, в том числе представители семейства Sclerosomatidae.
Во время вымирания в конце триаса вымерло 46–72% всех морских родов. Последствия вымирания в конце триаса были наибольшими в тропических широтах и более серьезными в Панталассе, чем в Тетисе или Северном океане. Экосистемы тропических рифов рухнули во время этого события и не восстановятся полностью до гораздо более позднего периода юрского периода. Сидячие питатели-фильтры и фотосимбиотические организмы пострадали в наибольшей степени.
Снизившись на границе триаса и юры, рифы существенно расширились в течение поздней юры, включая как губчатые рифы, так и коралловые рифы склерактиний. Позднеюрские рифы были похожи по форме на современные рифы, но имели больше микробных карбонатов и гиперкальцифицированных губок и имели слабое биогенное связывание. Рифы резко сократились в конце юрского периода, что вызвало соответствующее сокращение разнообразия десятиногих ракообразных. Самые ранние планктонные фораминиферы, Globigerinina, известны с конца ранней юры (середина тоара) западного Тетиса, распространились по всему Тетису к средней юре и стали глобально распространяться в тропических широтах к поздней юре. Кокколитофориды и динофлагелляты, впервые появившиеся в триасе, излучали излучение в период от ранней до средней юры, став заметными членами фитопланктона. Микроконхидные трубчатые черви, последний оставшийся отряд Tentaculita, группы животных с неопределенным родством, которые сходились с трубчатыми червями Spirorbis, были редки после триасового периода и были сведены к единственному роду Punctaconchus, который вымер в позднем Батоне. Самая старая из известных диатомовых водорослей - это позднеюрский янтарь из Таиланда, относящийся к живому роду Hemiaulus.
Морские лилии диверсифицировались на протяжении юрского периода, достигнув пика мезозойского разнообразия в поздней юре, в основном за счет излучения сидячих форм, принадлежащих отрядам Cyrtocrinida и Millericrinida. Начиная с ранней юры, ежи (морские ежи) претерпели существенную диверсификацию, в первую очередь за счет излучения нерегулярных (асимметричных) форм, которые приспосабливались к питанию отложениями. Темпы диверсификации резко упали в поздней юре.
 Eryon, polychelidan десятиногих ракообразные из поздней юры Германии.
Eryon, polychelidan десятиногих ракообразные из поздней юры Германии. Юрский период был важным временем для эволюции декапод. Первые настоящие крабы ( Brachyura ) известны с ранней юры, самые ранние из них - Eocarcinus praecursor из раннего плинсбаха в Англии, у которого отсутствовала крабоподобная морфология ( карцинизация ) современных крабов, и Eoprosopon klugi из позднего плинсбаха в Германии., которые могут принадлежать к живому семейству Homolodromiidae. Большинство крабов юрского периода известны только по частям панциря, что затрудняет определение их взаимоотношений. В то время как в ранней и средней юре крабы были редкостью, в поздней юре крабы стали многочисленными, поскольку они расширились из своей исконной среды обитания на илистом морском дне в места обитания с твердым субстратом, такие как рифы, с расщелинами на рифах, обеспечивающими убежище от хищников. Раки-отшельники также впервые появились в юрском периоде, и самым ранним из них является Schobertella hoelderi из позднего геттангского периода в Германии. Ранние раки-отшельники больше ассоциируются с раковинами аммонитов, чем с брюхоногими моллюсками. Глифеиды, которые сегодня известны только по двум видам, достигли пика своего разнообразия в юрский период: около 150 видов из общей летописи окаменелостей 250, известных с того периода. Юрские ракообразные отличались низким разнообразием по сравнению с современными, но известно несколько важных эволюционных инноваций, включая первые появления кальцитовых панцирных форм и видов с эпипланктонным образом жизни.
Разнообразие брахиопод уменьшилось во время триасово-юрского вымирания. Группы, несущие шпильки (Spiriferinida и Athyridida), сократились на границе триаса и юры и не восстановили свое биоразнообразие, вымерли в TOAE. Rhynchonellida и Terebratulida также уменьшились во время триасово-юрского вымирания, но восстановились в течение ранней юры; ни одна из кладов не претерпела больших морфологических изменений. В поздней юре брахиоподы существенно сократились; причины плохо изучены. Предлагаемые причины включают усиление хищничества, конкуренцию с двустворчатыми моллюсками, усиление биотурбации или повышенное давление пастбищ.
Аммониты были опустошены в конце триасового периода вымирания, и только несколько родов, принадлежащих к семейству Psiloceratidae подотряда Phylloceratina, выжили и стали предками всех более поздних юрских и меловых аммонитов. Аммониты резко разнообразились в течение ранней юры: отряды Psiloceratina, Ammonitina, Lytoceratina, Haploceratina, Perisphinctina и Ancyloceratina появились в юрском периоде. Фауны аммонитов в юрский период были региональными и были разделены примерно на 20 различных провинций и субпровинций в двух областях: северная высокоширотная Пан-Бореальная область, состоящая из Арктики, северной Панталассы и северной Атлантики, а также экваториально-южная Пан-Тетийская область. царство, в которое входили Тетис и большая часть Панталасса.
Вымирание в конце триаса оказало серьезное влияние на разнообразие двустворчатых моллюсков, хотя и мало повлияло на экологическое разнообразие двустворчатых моллюсков. Вымирание было избирательным, оказав меньшее влияние на глубоких нор, но нет доказательств различий в воздействии между живущими на поверхности (эпифауна) и роющими (инфауна) двустворчатыми моллюсками. Разнообразие на уровне семейства двустворчатых моллюсков после ранней юры было статичным, хотя родовое разнообразие постепенно увеличивалось на протяжении всего периода. Рудисты, доминирующие рифообразующие организмы мелового периода, впервые появились в поздней юре (середине оксфорда) на северной окраине западной части Тетиса, а к концу юры распространились до восточной части Тетиса.
Самые старые окончательные сведения о кальмароподобных белемнитах относятся к ранней юре (геттанг-синемурийский период) Европы и Японии; они распространились по всему миру в юрский период. Белемниты были обитателями мелководья, населявшими верхние 200 метров водной толщи на континентальных шельфах и в прибрежной зоне. Они были ключевыми компонентами юрских экосистем как хищники, так и жертвы, о чем свидетельствует обилие белемнитовых охранников в юрских породах.
Самые ранние вампироморфы, единственным живым членом которых является кальмар-вампир, впервые появились в раннем юрском периоде. Самые ранние осьминоги появились в средней юре, отделившись от своих ближайших живых родственников, вампироморфов, в период от триаса до ранней юры. Все юрские осьминоги известны исключительно по твердому гладиусу. Proteroctopus из позднего среднего юрского периода La Voulte-sur-Rhône lagerstätte, ранее считавшийся ранним осьминогом, теперь считается базальным таксоном за пределами клады, содержащей вампироморфов и осьминогов. Осьминоги, вероятно, произошли от донных ( придонных ) предков, которые жили на мелководье.