
Войти
| Цератозавр. Временной диапазон: Поздний юрский период, 153–148 Ma PreꞒ Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
| Слепок цератозавра из Кливлендского карьера Ллойда, выставленный в Музее естественной истории штата Юта | |
| Научная классификация | |
| Домен : | Eukaryota |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Семейство: | †Ceratosauridae |
| Род: | †Ceratosaurus. Marsh, 1884 |
| Типовой вид | |
| † Ceratosaurus nasicornis . Marsh, 1884 | |
| Другой виды | |
| |
| Синонимы | |
| |
Ceratosaurus (от греческого κέρας κέρατος, keras keratos, что означает «рог» и σαῦρος sauros, что означает «ящерица») был плотоядным тероподом динозавром в позднеюрском периоде (от кимериджа до титона ). Этот род был впервые описан в 1884 году американским палеонтологом Отниэлем Чарльзом Маршем на основе почти полного скелета, обнаруженного в Гарден-парке, Колорадо, в породах, принадлежащих Формация Моррисон. типовым видом является Ceratosaurus nasicornis.
Образец из Гарден-Парк остается наиболее полным скелетом, известным из этого рода, и с тех пор было описано лишь несколько дополнительных образцов. Два дополнительных вида, Ceratosaurus dentisulcatus и Ceratosaurus magnicornis, были описаны в 2000 году из двух фрагментарных скелетов из карьера Кливленд-Ллойд в Юте и из окрестностей Фруита, штат Колорадо.. Однако достоверность этих дополнительных видов была поставлена под сомнение, и все три скелета, возможно, представляют разные стадии роста одного и того же вида. В 1999 г. было сообщено об обнаружении первого экземпляра молоди. С 2000 г. был обнаружен и описан частичный образец из формации Лоуринья в Португалии, что свидетельствует о присутствии этого рода за пределами Северной Америки. О фрагментарных останках также сообщалось из Танзании, Уругвая и Швейцарии, хотя их отнесение к цератозавру в настоящее время не принято большинством палеонтологов.
Цератозавр был тероподом среднего размера. Исходный образец оценивается в 5,3 м (17 футов) или 5,69 м (18,7 фута) в длину, в то время как экземпляр, описанный как C. dentisulcatus, был больше, примерно 7 м (23 фута) в длину. Цератозавр характеризовался глубокими челюстями, которые поддерживали пропорционально очень длинные, похожие на лезвия зубы, выступающий гребневидный рог на средней линии морды и пару рогов над глазами. Передние конечности были очень короткими, но оставались полностью функциональными; на руке было четыре пальца. Хвост был глубоким сверху вниз. Ряд небольших остеодерм (кожных костей) присутствовал посередине шеи, спины и хвоста. Дополнительные остеодермы присутствовали в неизвестных местах на теле животного.
Цератозавр дал свое название Ceratosauria, кладе динозавров-теропод, которые рано отклонились от эволюционной линии, ведущей к современным птицам. В пределах Ceratosauria некоторые палеонтологи предположили, что это наиболее тесно связано с Genyodectes из Аргентины, у которого общие зубы сильно вытянутые. Более древний в геологическом отношении род Proceratosaurus из Англии, хотя первоначально описывался как предполагаемый предшественник Ceratosaurus, позже было обнаружено, что они не связаны между собой. Цератозавр разделял свою среду обитания с другими крупными родами теропод, включая Torvosaurus и Allosaurus, и было высказано предположение, что эти тероподы занимали разные экологические ниши, чтобы уменьшить конкуренцию. Цератозавр, возможно, охотился на динозавров, питающихся растениями, хотя некоторые палеонтологи предположили, что он охотился на водную добычу, такую как рыба. Носовой рог, вероятно, не использовался в качестве оружия, как первоначально предлагал Марш, но, скорее всего, использовался исключительно для демонстрации.
 Размер двух экземпляров по сравнению с человеком, голотип Ceratosaurus nasicornis (USNM 4735) оранжевого цвета и более крупный образец (UMNH VP 5278) синим цветом
Размер двух экземпляров по сравнению с человеком, голотип Ceratosaurus nasicornis (USNM 4735) оранжевого цвета и более крупный образец (UMNH VP 5278) синим цветом Ceratosaurus следовал строению тела, типичному для крупных динозавров-теропод. двуногий, он двигался на мощных задних лапах, а его руки были уменьшены в размерах. Образец USNM 4735, первый обнаруженный скелет и голотип Ceratosaurus nasicornis, имел длину 5,3 м (17 футов) или 5,69 м (18,7 фута) в длину, согласно отдельным источникам. Было ли это животное полностью выращенным, неясно. Отниэль Чарльз Марш в 1884 году предположил, что этот экземпляр весил примерно половину веса современного Аллозавра. В более поздних отчетах это было пересмотрено до 418 кг (922 фунта), 524 кг (1155 фунтов) или 670 кг (1480 фунтов). Еще три скелета, обнаруженные во второй половине 20 века, были значительно больше. Первая из них, UMNH VP 5278, была неофициально оценена Джеймсом Мэдсеном в длину около 8,8 м (29 футов), но позже была оценена в 7 м (23 фута) в длину. Его вес был рассчитан на 980 кг (2160 фунтов), 452 кг (996 фунтов) и 700 кг (1540 фунтов) в отдельных работах. Второй скелет, MWC 1, был несколько меньше UMNH VP 5278 и мог весить 275 кг (606 фунтов). Третий, еще не описанный, образец BYUVP 12893 был заявлен как самый крупный из обнаруженных, хотя оценки не были опубликованы. Другой экземпляр (ML 352), обнаруженный в Португалии в 2000 году, оценивался в длину 6 м (20 футов) и вес в 600 кг (1320 фунтов).
 Художественный оттиск C. nasicornis
Художественный оттиск C. nasicornis Точное количество позвонки неизвестны из-за нескольких щелей в позвоночнике голотипа Ceratosaurus nasicornis. Не менее 20 позвонков образовали шею и заднюю часть перед крестцом. В средней части шеи centra (тела) позвонков были такой же длины, как и их высота, в то время как в передней и задней частях шеи центральные части были короче своего роста. Выступающие вверх нервные отростки были сравнительно большими, а в спинных (задних) позвонках были такой же высоты, как и центры позвонков. Крестец, состоящий из шести сросшихся крестцовых позвонков , выгнут вверх, при этом его позвоночные центры сильно уменьшены по высоте в средней части, как это имеет место у некоторых других цератозавров.. Хвост включает примерно 50 хвостовых позвонков и составляет примерно половину общей длины животного; на голотипе он оценивается в 2,84 м (9,33 фута). Хвост был глубоким сверху вниз из-за высоких нервных шипов и удлиненных шевронов, костей, расположенных ниже тел позвонков. Как и у других динозавров, он уравновешивал тело и содержал массивную caudofemoralis мышцу, которая отвечала за толчок вперед во время передвижения, оттягивая верхнюю часть бедра назад при сокращении.
 . Отличительные особенности черепа Ceratosaurus: Сращенные левая и правая носовые кости образуют выдающийся носовой рог (вверху), а зубы верхней челюсти исключительно длинные (внизу). Эти окаменелости являются частью образца MWC 1 из Фруита, Колорадо и выставлены в местном музее путешествий динозавров.
. Отличительные особенности черепа Ceratosaurus: Сращенные левая и правая носовые кости образуют выдающийся носовой рог (вверху), а зубы верхней челюсти исключительно длинные (внизу). Эти окаменелости являются частью образца MWC 1 из Фруита, Колорадо и выставлены в местном музее путешествий динозавров. лопатка (лопатка) была слита с коракоид, образующий единую кость без какой-либо видимой границы между двумя исходными элементами. Голотип C. nasicornis был обнаружен с сочлененной левой передней конечностью, включая неполный manus (рука). Хотя отделил во время подготовки, заранее была сделана слепка окаменелости, чтобы задокументировать исходное относительное положение костей. Кости запястья не были известны ни по одному экземпляру, поэтому некоторые авторы предполагают, что они были потеряны в этом роду. В статье 2016 года Мэтью Каррано и Джона Чуаньер предположили, что, вероятно, присутствовали один или несколько хрящевых (не костных) запястных костей, о чем свидетельствует зазор между костями предплечья и пястными костей, а также текстура поверхности в пределах этого зазора видна в отливке. В отличие от большинства более производных от тероподов, у которых на каждой кисти было только три пальца (цифры I – III), у Ceratosaurus сохранились четыре пальца, а палец IV уменьшился в размере. Первая и четвертая пястные кости были короткими, а вторая была немного длиннее третьей. Пястные кости и особенно первые фаланги были пропорционально очень короткими, в отличие от большинства других базальных теропод. На голотипе сохранились только первые фаланги пальцев II, III и IV; общее количество фаланг и копытных (когтей) неизвестно. Анатомия I пястной кости указывает на то, что изначально фаланги присутствовали и на этом пальце. pes (ступня) состояла из трех опорных пальцев, пронумерованных II – IV. Цифра I, которая у теропод обычно сокращается до росового когтя, не касающегося земли, в голотипе не сохранилась. Марш в своем первоначальном описании 1884 года предположил, что этот палец был утерян у Ceratosaurus, но Чарльз Гилмор в своей монографии 1920 года отметил область прикрепления на второй плюсневой кости, демонстрирующую присутствие этот палец.
Цератозавр, уникально среди теропод, обладал небольшими, удлиненными и неправильно сформированными остеодермами (кожными костями) вдоль средней линии тела. Такие остеодермы были обнаружены над нервными отростками шейных позвонков 4 и 5, а также хвостовых позвонков с 4 по 10 и, вероятно, образовывали непрерывный ряд, который мог тянуться от основания черепа до большей части хвоста. Как предположил Гилмор в 1920 году, их положение в матрице горных пород, вероятно, отражает их точное положение в живом животном. Было обнаружено, что остеодермы над хвостом отделены от нервных шипов на 25 мм (0,98 дюйма) до 38 мм (1,5 дюйма), что, возможно, составляет кожу и мышцы, присутствующие между ними, в то время как мышцы шеи были гораздо ближе к нервным шипам. Помимо средней линии тела, кожа содержала дополнительные остеодермы, на что указывает крупная четырехугольная пластинка размером 58 мм (2,3 дюйма) на 70 мм (2,8 дюйма), обнаруженная вместе с голотипом; положение этой пластины на теле неизвестно. Образец UMNH VP 5278 был также обнаружен с рядом остеодерм, которые были описаны как аморфные по форме. Хотя большинство из этих костей были обнаружены на расстоянии не более 5 м от скелета, они не были напрямую связаны с какими-либо позвонками, в отличие от голотипа C. nasicornis, поэтому их исходное положение на тело не может быть выведено из этого экземпляра.
 Схема голотипа черепа Ceratosaurus nasicornis сверху и сбоку, сделанная Чарльзом Гилмором, 1920 г.: Эта реконструкция теперь считается слишком широкий на виде сверху.
Схема голотипа черепа Ceratosaurus nasicornis сверху и сбоку, сделанная Чарльзом Гилмором, 1920 г.: Эта реконструкция теперь считается слишком широкий на виде сверху. череп был довольно большим по сравнению с остальной частью его тела. У голотипа C. nasicornis он имеет длину 55 см (22 дюйма), измеренную от кончика морды до затылочного мыщелка , который соединяется с первым шейным позвонком. Ширину этого черепа трудно реконструировать, так как он сильно искажен, а реконструкция Гилмора 1920 года была позже признана слишком широкой. Довольно полный череп образца MWC 1, по оценкам, имел длину 60 см (24 дюйма) и ширину 16 см (6,3 дюйма); этот череп был несколько более вытянутым, чем у голотипа. Задняя часть черепа была построена легче, чем у некоторых других более крупных теропод, из-за обширных отверстий в черепе, но челюсти были глубокими, чтобы поддерживать пропорционально большие зубы. слезная кость образовывала не только задний край анторбитального отверстия, большое отверстие между глазом и костной ноздрей , но и часть его верхнего края, в отличие от членов родственного Abelisauridae. Квадратная кость , которая была соединена с нижней челюстью на ее нижнем конце для образования челюстного сустава, была наклонена так, что челюстной сустав был смещен назад по отношению к затылочному мыщелку. Это также привело к расширению основания бокового височного отверстия, большого отверстия за глазами.
Самой отличительной чертой был выступающий рог, расположенный на средней линии черепа за костями. ноздри, образовавшиеся из сросшихся бугорков левой и правой носовых костей. Из окаменелостей известно только костяное ядро рога - у живого животного оно могло поддерживать ороговевшую оболочку. В то время как основание рогового ядра было гладким, его верхние две трети были морщинистыми и выстланы бороздками, которые при жизни могли бы содержать кровеносные сосуды. В голотипе ядро рога имеет длину 13 см (5,1 дюйма) и ширину 2 см (0,79 дюйма) у основания, но быстро сужается до 1,2 см (0,47 дюйма) дальше вверх; его высота составляет 7 см (2,8 дюйма). В черепе MWC 1 он длиннее и ниже. У живого животного рог, вероятно, был бы более удлиненным из-за своей ороговевшей оболочки. За носовым рогом носовые кости образовывали овальную бороздку; и эта бороздка, и носовой рог служат признаками, чтобы отличить Ceratosaurus от родственных родов. Помимо большого носового рога, цератозавр обладал меньшими полукруглыми костными выступами перед каждым глазом, подобными таковым у аллозавра. Эти гребни образованы слезными костями. У молодых особей все три рога были меньше, чем у взрослых, и две половины сердцевины носового рога еще не срослись.
предчелюстные кости, образующие кончик морды, имел всего по три зуба с каждой стороны, меньше, чем у большинства других теропод. На голотипе верхнечелюстные кости верхней челюсти были выровнены по 15 лезвийных зубов с каждой стороны. Первые восемь из этих зубов были очень длинными и крепкими, но начиная с девятого зуба они постепенно уменьшались в размерах. Как типично для теропод, они имели мелкие зубчатые края, которые в голотипе содержали примерно 10 зубцов на 5 мм (0,20 дюйма). Образец MWC 1 показал всего от 11 до 12, а образец UMNH VP 5278 - 12 зубов в каждой верхней челюсти; у последнего экземпляра зубы были более прочными и более изогнутыми. У всех образцов коронки верхней челюсти были исключительно длинными. В образце УМНХ ВП 5278 они имели длину до 9,3 см (3,7 дюйма), что равно минимальной высоте нижней челюсти. На голотипе они имеют длину 7 см (2,8 дюйма), что даже превышает минимальную высоту нижней челюсти. У других теропод сравнимая длина зубов известна только по возможно близкородственному Genyodectes. Напротив, некоторые представители Abelisauridae имеют очень короткие коронки зубов. На голотипе каждая половина зубной , зубной кости нижней челюсти , была оснащена 15 зубами, которые однако плохо сохранились. Оба образца MWC 1 и UMNH VP 5278 показывают только 11 зубов в каждой зубной части, которые, как показывает последний образец, были немного более прямыми и менее прочными, чем у верхней челюсти.
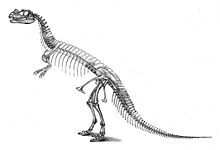 C. nasicornis, реставрация скелета Отниэлем Чарльзом Маршем из 1892 года, изображенная в неправильном вертикальном положении и с избыточными позвонками в позвоночнике, приводящими к чрезмерно удлиненному стволу
C. nasicornis, реставрация скелета Отниэлем Чарльзом Маршем из 1892 года, изображенная в неправильном вертикальном положении и с избыточными позвонками в позвоночнике, приводящими к чрезмерно удлиненному стволу Первый образец, голотип USNM 4735, был обнаружен и раскопан фермером Маршаллом Паркером Фельчем в 1883 и 1884 годах. Найденный в сочленении, с костями, все еще соединенными друг с другом, он был почти готов, включая череп. Значительные недостающие части включают неизвестное количество позвонков; все ребра туловища, кроме последних; плечевые кости (кости плеча); дистальные кости пальцев обеих рук; большая часть правой передней конечности; большая часть левой задней конечности; и большая часть ног. Образец был найден заключенным в твердый песчаник; череп и позвоночник были сильно искажены во время окаменелости. Место открытия, расположенное в районе Гарден Парк к северу от Каньон-Сити, Колорадо, и известное как карьер Фелч 1, считается одним из самых богатых ископаемых памятников Формация Моррисон. Многочисленные окаменелости динозавров были обнаружены в этом карьере еще до открытия цератозавра, в первую очередь почти полный образец аллозавра (USNM 4734) в 1883 и 1884 годах.
После раскопок образец был отправлен на Музей естественной истории Пибоди в Нью-Хейвене, где он был изучен Маршем, который описал его как новый род и вид Ceratosaurus nasicornis в 1884 году. Имя Ceratosaurus может быть переведено как «рог ящерицы». "(от греческого κερας / κερατος, keras / keratos -« рог »и σαυρος / sauros -« ящерица ») и nasicornis с« носовым рогом »(от латинского nasus -« нос »и рога -« рог »). Учитывая полноту экземпляра, недавно описанный род был в то время самым известным тероподом, обнаруженным в Америке. В 1898 и 1899 годах образец был передан в Национальный музей естественной истории в Вашингтоне, округ Колумбия, вместе со многими другими окаменелостями, первоначально описанными Маршем. Только часть этого материала была полностью подготовлена, когда прибыла в Вашингтон; последующая подготовка длилась с 1911 по конец 1918 года. Упаковка и доставка из Нью-Хейвена в Вашингтон нанесли некоторый ущерб экземпляру Ceratosaurus. В 1920 году Чарльз Гилмор опубликовал подробное переописание этого и других образцов теропод, полученных из Нью-Хейвена, включая почти полный экземпляр аллозавра, извлеченный из того же карьера.
 Реконструкция голотипа экземпляра C. nasicornis от Чарльза У. Гилмора, 1920, показаны известные элементы
Реконструкция голотипа экземпляра C. nasicornis от Чарльза У. Гилмора, 1920, показаны известные элементы В статье 1892 года Марш опубликовал первую реконструкцию скелета Ceratosaurus, которая изображает животное длиной 22 фута (6,7 м) и длиной 12 м. футов (3,7 м) в высоту. Как отмечал Гилмор в 1920 году, туловище в этой реконструкции было изображено слишком длинным, включающим по крайней мере шесть спинных позвонков. Эта ошибка повторялась в нескольких последующих публикациях, включая первую реконструкцию жизни, которая была нарисована в 1899 году Фрэнком Бондом под руководством Чарльза Р. Найта, но опубликована только в 1920 году. Более точная реконструкция жизни, опубликованный в 1901 г., был подготовлен Джозефом М. Глисоном, снова под руководством Найта. Голотип был установлен Гилмором в 1910 и 1911 годах, и с тех пор выставлялся в Национальном музее естественной истории. Большинство ранних реконструкций показывают цератозавра в вертикальном положении, волоча хвостом по земле. Гилмор на голотипе, напротив, опередил свое время: вдохновленный костями верхней части бедра, которые были обнаружены под углом к голени, он изобразил животное как бегущее животное с горизонтальной, а не вертикальной позой и хвостом, который не касался земли. Из-за того, что окаменелости сильно уплощены, Гилмор установил образец не как отдельно стоящий скелет, а как барельеф внутри искусственной стены. Поскольку кости были частично встроены в бляшку, доступ к исследованиям был ограничен. В ходе обновления экспозиции динозавров в музее в период с 2014 по 2019 год экземпляр был демонтирован и освобожден от обшивки. На новой выставке, которая откроется в 2019 году, монтировку планируется заменить отдельно стоящей, а оригинальные кости будут храниться в коллекции музея, чтобы обеспечить полный доступ для ученых.
 первая реставрация жизни цератозавра, нарисованная в 1899 году Фрэнком Бондом под руководством Чарльза Р. Найта, но не опубликованная до 1920 года
первая реставрация жизни цератозавра, нарисованная в 1899 году Фрэнком Бондом под руководством Чарльза Р. Найта, но не опубликованная до 1920 года После открытия голотипа C. nasicornis, значительная находка цератозавра не была сделана до начала 1960-х годов, когда палеонтолог Джеймс Мэдсен и его команда обнаружили фрагментарный, разрозненный скелет, включая череп (UMNH VP 5278), в карьере динозавров Кливленд-Ллойд в штате Юта. Эта находка представляет собой один из самых крупных известных экземпляров цератозавра. Второй сочлененный образец, включающий череп (MWC 1), был обнаружен в 1976 году Тором Эриксоном, сыном палеонтолога Ланса Эриксона, недалеко от Фруита, Колорадо. Достаточно полный экземпляр, без нижней челюсти, предплечий и гастралии. Череп, хотя и довольно полный, был обнаружен разрозненным и сильно уплощенным с боков. Хотя он был крупным, он еще не достиг взрослого размера, о чем свидетельствуют открытые швы между костями черепа. Научно точные трехмерные реконструкции черепа для использования в музейных экспонатах были произведены с использованием сложного процесса, включающего формование и отливку отдельных оригинальных костей, исправление деформаций, реконструкцию недостающих частей, сборку слепков костей в их правильное положение и
Образцы Фруита и Кливленд-Ллойд были описаны Мэдсеном и Сэмюэлем Полом Уэллсом в монографии 2000 года, при этом образец из Юты был отнесен к новый вид C. dentisulcatus и образец из Колорадо - новый вид C. magnicornis. Название dentisulcatus относится к параллельным бороздкам, присутствующим на внутренней стороне предчелюстных зубов и первых трех зубов нижней челюсти в этом образце; magnicornis указывает на больший носовой рог. Однако валидность обоих видов подвергалась сомнению в последующих публикациях. Брукс Бритт и его коллеги в 2000 году заявили, что голотип C. nasicornis на самом деле был ювенильной особью, а два более крупных вида представляли взрослую особь одного вида. Оливер Раухут в 2003 году, а также Мэтью Каррано и Скотт Сэмпсон в 2008 году рассмотрели анатомические различия, приведенные Мэдсеном и Уэллсом, чтобы подтвердить, что эти дополнительные виды представляют онтогенетические (связанные с возрастом) или индивидуальные вариации.
 Частичный ювенильный образец, Североамериканский музей древней жизни
Частичный ювенильный образец, Североамериканский музей древней жизни Еще один образец (BYUVP 12893) был обнаружен в 1992 году в карьере Агатового бассейна к юго-востоку от Мура, штат Юта, но все еще ожидает описания. Образец, который считается самым крупным из представителей этого рода, включает переднюю половину черепа, семь фрагментов тазовых спинных позвонков, а также сочлененный таз и крестец. В 1999 году Бритт сообщил об открытии скелета цератозавра, принадлежащего молодому человеку. Обнаруженный в Bone Cabin Quarry в Вайоминге, он на 34% меньше голотипа C. nasicornis и состоит из целого черепа, а также 30% остальной части скелета, включая полный таз.
Помимо этих пяти находок скелетов, фрагментарные останки цератозавра были зарегистрированы в различных местах стратиграфических зон 2 и 4-6 формации Моррисон, включая некоторые из основных мест окаменелости формации. Национальный памятник динозавров, штат Юта, обнаружил изолированную правую предчелюстную кость (номер экземпляра DNM 972); большая лопатка (скапулокоракоид) была обнаружена в Комо Блафф в Вайоминге. Другой образец происходит из Dry Mesa Quarry, Колорадо, и включает левый скапулокоракоид, а также фрагменты позвонков и костей конечностей. В карьере Майгатт Мур, штат Колорадо, этот род известен по зубам.
С 1909 по 1913 год немецкие экспедиции Берлина музея für Naturkunde обнаружил разнообразную фауну динозавров из формации Tendaguru в Германской Восточной Африке, на территории нынешней Танзании. Хотя обычно они считаются наиболее важным местом обитания африканских динозавров, крупные динозавры-тероподы известны лишь по немногим и очень фрагментарным останкам. В 1920 году немецкий палеонтолог Вернер Яненш отнес несколько спинных позвонков из карьера "TL" к Ceratosaurus, как Ceratosaurus sp. (неопределенных видов). В 1925 году Яненш назвал новый вид Ceratosaurus, C. roechlingi, на основе фрагментарных останков из карьера «Mw», включающих квадратную кость, малоберцовую кость, фрагменты хвостовых позвонков и другие фрагменты. Этот экземпляр произошел от особи, значительно крупнее голотипа C. nasicornis.
 Реставрация C. nasicornis, выполненная Джозефом М. Глисоном с 1901 года, выполненная под руководством Чарльза Р. Найта.
Реставрация C. nasicornis, выполненная Джозефом М. Глисоном с 1901 года, выполненная под руководством Чарльза Р. Найта.В своей монографии 2000 года Мэдсен и Уэллс подтвердили принадлежность этих находок к цератозаврам. Кроме того, они отнесли несколько зубов к роду, который первоначально был описан Яненшем как возможный вид Labrosaurus, Labrosaurus (?) Stechowi. Другие авторы поставили под сомнение принадлежность каких-либо находок Тендагуру к Ceratosaurus, отметив, что ни один из этих образцов не имеет признаков, диагностических для этого рода. В 2011 году Раухут обнаружил, что C. roechlingi и Labrosaurus (?) Stechowi являются возможными цератозавридами, но обнаружил, что они не являются диагностическими на уровне рода, и поэтому обозначил их как nomina dubia (сомнительные названия). В 1990 году Тимоти Роу и Жак Готье упомянули еще один вид Ceratosaurus из Тендагуру, Ceratosaurus ingens, который якобы был воздвигнут Яненшем в 1920 году и основывался на 25 отдельных очень больших зубах до 15 см (5,9 дюйма).) в длину. Однако Яненш отнес этот вид к мегалозаврам, а не к цератозаврам; следовательно, это имя может быть простой ошибкой копирования. Раухут в 2011 году показал, что Megalosaurus ingens не был тесно связан ни с Megalosaurus, ни с Ceratosaurus, но, возможно, представляет собой кархародонтозаврид, а вместо этого.
В 2000 и 2006 годах палеонтологи во главе с Октавио Матеус описал находку из формации Лоринья в центрально-западной Португалии (ML 352) как новый экземпляр Ceratosaurus, состоящий из правого бедренной кости (верхняя кость бедра), левая большеберцовая кость (большеберцовая кость) и несколько изолированных зубов, найденных на скалах пляжа Валмитао, между муниципалитетами Лоринья и Торрес Ведрас. Кости были обнаружены в мелкозернистых песчаниках от желтого до коричневого цвета, которые были отложены реками в виде пойменных отложений и относятся к нижним уровням пачки Порто-Ново, возраст которой считается поздним кимериджским.. Дополнительные кости этого человека (SHN (JJS) -65), включая левую бедренную кость, правую большеберцовую кость и частично левую малоберцовую кость (телячья кость), с тех пор были обнажены из-за прогрессирующей эрозии утеса.. Хотя изначально эти дополнительные элементы были частью частной коллекции, эти дополнительные элементы стали официально храниться после того, как частная коллекция была передана Sociedade de História Natural в Торрес-Ведрас, и были подробно описаны в 2015 году. Образец был отнесен к виду Ceratosaurus dentisulcatus Матеусом и коллегами в 2006 г. Обзор 2008 г., проведенный Каррано и Сэмпсоном, подтвердил отнесение к Ceratosaurus, но пришел к выводу, что отнесение к какому-либо конкретному виду в настоящее время невозможно. В 2015 году Элизабет Малафаиа и его коллеги, поставившие под сомнение достоверность C. dentisulcatus, отнесли образец к Ceratosaurus aff. Ceratosaurus nasicornis.
В других сообщениях упоминается об одном зубе, найденном в Мутье, Швейцария. Первоначально названный Яненшем в 1920 году как Labrosaurus meriani, зуб позже был назначен Ceratosaurus sp. (неизвестного вида) Мэдсена и Уэллса. В 2008 году Матиас Сото и Даниэль Переа описали зубы из формации Такуарембо в Уругвае, включая предполагаемую коронку предчелюстной кости. На его внутренней стороне видны вертикальные бороздки, а на переднем крае отсутствуют зубчики; эти особенности в этой комбинации известны только от Ceratosaurus. Авторы, однако, подчеркивают, что отнесение европейского и африканского материала к цератозавру невозможно, так как останков мало, и, кроме того, отмечают, что отнесение европейского и африканского материала к цератозавру следует рассматривать с осторожностью.
 С. nasicornis голотип, установленный Чарльзом Гилмором в 1910 и 1911 годах, Национальный музей естественной истории
С. nasicornis голотип, установленный Чарльзом Гилмором в 1910 и 1911 годах, Национальный музей естественной истории В своем первоначальном описании голотипа Ceratosaurus nasicornis и последующих публикациях Марш отметил ряд характеристики, которые не были известны у всех других известных в то время теропод. Две из этих черт, сросшийся таз и сросшийся плюсневой кость, были известны у современных птиц и, по словам Марша, ясно демонстрируют тесную связь между последними и динозаврами. Чтобы отделить этот род от Allosaurus, Megalosaurus и целурозавров, Марш сделал Ceratosaurus единственным членом как нового семейства, и Ceratosauridae, и нового инфраотряд, Ceratosauria. Это было поставлено под сомнение в 1892 году Эдвардом Дринкером Коупом, соперником Марша в Bone Wars, который утверждал, что такие отличительные особенности, как носовой рог, просто показывают, что C. nasicornis был отдельным видом, но этого было недостаточно, чтобы оправдать отдельный род. Следовательно, он отнес C. nasicornis к роду Megalosaurus, создав новую комбинацию Megalosaurus nasicornis.
Хотя Ceratosaurus сохранялся как отдельный род во всех последующих анализах, его отношения оставались спорными в течение следующего столетия. И Ceratosauridae, и Ceratosauria не получили широкого признания, были идентифицированы лишь несколько и малоизвестных дополнительных членов. На протяжении многих лет отдельные авторы классифицировали Ceratosaurus в пределах Deinodontidae, Megalosauridae, Coelurosauria, Carnosauria и Deinodontoidea. В своей редакции 1920 года Гилмор утверждал, что этот род был наиболее базальным тероподом, известным после триасового периода, поэтому не был тесно связан с каким-либо другим современным тероподом, известным в то время; таким образом, он имеет собственное семейство Ceratosauridae. Однако только после создания кладистического анализа в 1980-х годах первоначальное заявление Марша о Ceratosauria как об отдельной группе получило распространение. В 1985 году было обнаружено, что недавно открытые южноамериканские роды Abelisaurus и Carnotaurus были тесно связаны с Ceratosaurus. В 1986 году Готье признал, что Coelophysoidea имеет близкое родство с Ceratosaurus, хотя согласно последним исследованиям эта клада выходит за рамки Ceratosauria. С тех пор было обнаружено множество дополнительных представителей Ceratosauria.
Ceratosauria рано отделилась от эволюционной линии, ведущей к современным птицам, поэтому считается базальной внутри теропод. Сама цератозаврия содержит группу производных (небазальных) членов семейств Noasauridae и Abelisauridae, которые заключены в скобки внутри клады Abelisauroida, а также ряд базальные члены, такие как Elaphrosaurus, Deltadromeus и Ceratosaurus. Положение Ceratosaurus среди базальных цератозавров обсуждается. Некоторые анализы рассматривали Ceratosaurus как наиболее производных от базальных представителей, образующих сестринский таксон Abelisauroida. Оливер Раухут в 2004 году предложил Genyodectes в качестве сестринского таксона Ceratosaurus, поскольку оба рода характеризуются исключительно длинными зубами в верхней челюсти. Раухут сгруппировал Ceratosaurus и Genyodectes в семейство Ceratosauridae, за чем последовали несколько более поздних отчетов.
Шо Ван и его коллеги в 2017 году пришли к выводу, что Noasauridae не гнездились внутри Abelisauroida, как предполагалось ранее, а вместо этого были более базальный, чем Ceratosaurus. Поскольку ноазавриды использовались в качестве фиксированной точки для определения клад Abelisauroida и Abelisauridae, эти клады, следовательно, будут включать гораздо больше таксонов по определению, включая Ceratosaurus. В последующем исследовании 2018 года Рафаэль Делькур согласился с этими результатами, но указал, что, как следствие, Abelisauroida необходимо заменить на более старый синоним Ceratosauroida, который до сих пор использовался редко. Для Abelisauridae Делькур предложил новое определение, исключающее Ceratosaurus, что позволяет использовать название в его традиционном смысле. Ван и его коллеги также обнаружили, что Ceratosaurus и Genyodectes образуют кладу с аргентинским родом Eoabelisaurus. Делькур использовал название Ceratosauridae для обозначения этой же клады и предложил определить Ceratosauridae как содержащие все таксоны, которые более тесно связаны с Ceratosaurus, чем с абелизавридом Carnotaurus.
 Конный слепок ювенильного скелета, Музей открытия динозавров
Конный слепок ювенильного скелета, Музей открытия динозавров Следующая кладограмма, показывающая взаимоотношения цератозавров, основана на филогенетическом анализе, проведенном Диего Полом и Оливером Раухутом в 2012 г.:
| Ceratosauria |
|
 Сухой сезон в карьере Мигатт-Мур, где цератозавр (в центре) и
Сухой сезон в карьере Мигатт-Мур, где цератозавр (в центре) и  Реконструированные скелеты аллозавра и C. nasicornis в боевых позах, на выставке Dinoworld Expo, Йокогама
Реконструированные скелеты аллозавра и C. nasicornis в боевых позах, на выставке Dinoworld Expo, Йокогама  Восстановление кормления C. nasicornis
Восстановление кормления C. nasicornis  слепок руки C. nasicornis (
слепок руки C. nasicornis ( слепок черепа голотипа C. nasicornis, показывающий большой носовой рог,
слепок черепа голотипа C. nasicornis, показывающий большой носовой рог,  Скелеты C. nasicornis и
Скелеты C. nasicornis и