
Войти
| Мультитуберкулезные Временной диапазон: средняя юра - поздний эоцен. PreꞒ Ꞓ О S D C п Т J K Стр. N | |
|---|---|
 | |
| Скелет Катопсбаатара | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | Млекопитающие |
| Клэйд: | Тереобразные |
| Порядок: | † Multituberculata Cope, 1884 г. |
| Подзаказы | |
| |
Multituberculata (широко известные как multituberculata, названные в честь нескольких бугорков их зубов) - это вымерший отряд грызуновоподобных млекопитающих с окаменелостями, насчитывающими более 120 миллионов лет. Впервые они появились в средней юре и достигли пика разнообразия в меловом периоде. В конце концов, они пришли в упадок с позднего палеоцена, исчезнув из известных летописей окаменелостей в конце эоцена. Это самый разнообразный отряд мезозоя. млекопитающие с более чем 200 известными видами, от размера мыши до бобра. Эти виды занимали разнообразные экологические ниши, от обитания в норках до беличьего дендрария и личинок тушканчиков. Мультитуберкулятов обычно помещают в качестве коронных млекопитающих вне одной из двух основных групп живых млекопитающих - Theria, включая плацентарных и сумчатых, и Monotremata - но ближе к Theria, чем к монотремам. Тем не менее, по крайней мере, одно исследование обнаружило потенциальный статус сестринских таксонов монотрем / Australosphenida.
 Восстановление Taeniolabis, крупнейшего мультитуберкулеза при примерно 100 кг.
Восстановление Taeniolabis, крупнейшего мультитуберкулеза при примерно 100 кг. Мультитуберкуляты имели черепную и зубную анатомию, внешне похожую на грызунов, таких как мыши и крысы, с щечными зубами, отделенными от долотообразных передних зубов широкой беззубой щелью ( диастема ). Каждый щечный зуб имел несколько рядов маленьких бугорков (или бугорков, отсюда и название), которые действовали на аналогичные ряды в зубах челюсти; точная гомология этих куспидов териановым остаётся предметом споров. В отличие от грызунов, у которых постоянно растут зубы, у мультитуберкулезных особей протекали паттерны замены зубов, типичные для большинства млекопитающих (хотя, по крайней мере, у некоторых видов нижние резцы продолжали прорезаться еще долгое время после закрытия корня). Мультитуберкуляты отличаются наличием массивного четвертого нижнего премоляра, плагиалакоида ; другие млекопитающие, такие как Plesiadapiformes и дипротодонтовые сумчатые, также имеют аналогичные премоляры как на верхней, так и на нижней челюстях, но у мультитуберкулезных животных этот зуб массивный, и верхние премоляры не видоизменяются таким образом. У базальных мульти-туберкулезных животных все три нижних премоляра были плагиоулакоидами, увеличиваясь в размерах кзади, но у Cimolodonta остался только четвертый нижний премоляр, третий остался только в виде рудиментарного штифтового зуба, а в некоторых таксонах, таких как gondwanatherians и taeniolabidoideans, плагиаулакоид полностью исчез или был преобразован в коренной зуб.
 Череп Птилода. Обратите внимание на массивный лопаточный нижний премоляр.
Череп Птилода. Обратите внимание на массивный лопаточный нижний премоляр. В отличие от грызунов и подобных им терианов, у мультитуберкулезных челюстей был палинальный ход челюсти (спереди назад) вместо пропалинального (сзади вперед) или поперечного (из стороны в сторону); как следствие, их мускулатура челюстей и ориентация бугров радикально различаются. Палинальные удары челюстью почти полностью отсутствуют у современных млекопитающих (за исключением, возможно, дюгоня ), но также присутствуют у харамииданов, аргиролагоидов и тритилодонтидов, которые исторически объединялись с мультитуберкулезными на этом основании. Считается, что многотуберкулезное жевание происходит в двухтактном цикле: во-первых, пища, удерживаемая на месте последним верхним премоляром, разрезалась лопаточными нижними премолярами, когда зубной камень двигался ортально (вверх). Затем нижняя челюсть двигалась палинально, растирая пищу между рядами бугров коренных зубов.
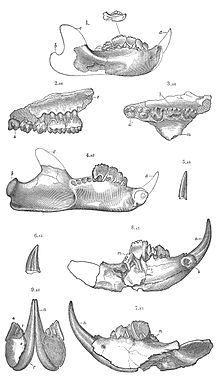 Нижняя челюсть и зубы аллодонтидных мультитуберкулезных
Нижняя челюсть и зубы аллодонтидных мультитуберкулезных Строение таза Multituberculata предполагает, что они родили крошечных беспомощных, недоразвитых детенышей, похожих на современных сумчатых, таких как кенгуру.
По крайней мере, две линии развивали гипсодонтию, при которой зубная эмаль выходит за пределы линии десен: lambdopsalid taeniolabidoideans и sudamericid gondwanatheres. Последние, существовавшие уже в меловом периоде, являются самой ранней известной линией пастбищных млекопитающих. Вид из геопарка «Лес динозавров Кацуяма» может предложить еще более ранний пример адаптации к травоядному, поскольку он датируется нижним меловым периодом примерно 120 миллионов лет назад.
Исследования, опубликованные в 2018 году, показали, что у мультитуберкулезных животных относительно сложный мозг, а некоторые области мозга даже отсутствуют у терианских млекопитающих.
Мультитуберкуляты впервые появляются в летописи окаменелостей в юрский период, а затем выжили и даже доминировали более ста миллионов лет, дольше, чем любой другой отряд млекопитающих, включая плацентарных млекопитающих. Самые ранние известные multituberculates взяты из средней юры ( бат \ 166-168 млн лет назад) из Англии и России, в том числе Hahnotherium и Kermackodon из лесного Marble Формирования Англии и Tashtykia и Tagaria из свиты Итат России. Эти формы известны только по изолированным зубам, которые имеют большое сходство с зубами эухарамиданов, которым они, как предполагается, имеют близкое родство.
В течение мелового периода мультитуберкуляты излучали широкий спектр морфотипов, в том числе беличеподобных древесных птилодонтов. Большинство видов multituberculata, по-видимому, были уничтожены во время события KT (вымирание динозавров), но они, похоже, были одними из первых, кто восстановился и снова диверсифицировался. Своеобразная форма их последнего нижнего премоляра - их самая выдающаяся особенность. Эти зубы были больше и более вытянуты, чем другие щечные зубы, и имели окклюзионную поверхность, образующую зубчатое режущее лезвие. Хотя можно предположить, что это использовалось для измельчения семян и орехов, считается, что большинство мелких мультитуберкулезных особей также дополняли свой рацион насекомыми, червями и фруктами. Следы зубов, приписываемые мультитуберкулезу, известны на окаменелостях Champsosaurus, что указывает на то, что по крайней мере некоторые из этих млекопитающих были падальщиками.
Птилодонтом, который процветал в Северной Америке, был Ptilodus. Благодаря хорошо сохранившимся экземплярам Ptilodus, найденным в бассейне Бигхорн, штат Вайоминг, мы знаем, что эти мультитуберкулезные животные были способны отводить и приводить свои большие пальцы ног, и, таким образом, их подвижность стопы была аналогична подвижности ног современных белок, которые спускаются с деревьев головой вперед..
 Восстановление Катопсбаатара
Восстановление Катопсбаатара В Европе другое семейство мультитуберкулезных животных было столь же успешным - Kogaionidae, впервые обнаруженное в Хацеге, Румыния. У них также развился увеличенный лопаточный нижний премоляр. Hainina, самый успешный род, изначально считался птиодонтом. Однако более подробный анализ этого рода выявил меньшее количество зубных бугров и оставшийся пятый премоляр - уникальное сочетание примитивных и продвинутых черт, указывающих на то, что Хайнина были родственниками некоторых юрских родов и что увеличенные лопаточные премоляры были приобретены независимо в Европа и Северная Америка.
Другая группа мультитуберкулятов, taeniolabids, была тяжелее и массивнее, что указывало на то, что они жили полностью земной жизнью. Самые крупные экземпляры весили около 100 кг, что делает их сопоставимыми по размеру с крупными грызунами, такими как Castoroides. Наибольшего разнообразия в Азии они достигли в конце мелового периода и палеоцена, что позволяет предположить, что они произошли оттуда.
Около 80 родов из многобугорчатых известны, в том числе Lambdopsalis, птилодус и Meniscoessus. В северном полушарии в конце мелового периода более половины типичных видов наземных млекопитающих были мультитуберкулезными. В то время как большинство млекопитающих - наряду с птицами, другими динозаврами и большинством других видов жизни - были уничтожены во время события KT (вымирание динозавров 65 миллионов лет назад), большая часть млекопитающих, которые появляются в летописи окаменелостей после вымирание - мультитуберкулезные.
Группа продолжала доминировать на суше в следующие двадцать миллионов лет палеоцена, достигнув пика разнообразия в эту эпоху. Плацентарное разнообразие тем временем было ограничено; только после того, как количество туберкулезных групп уменьшится после датского периода, количество плацентарных млекопитающих резко увеличится.
 Восстановление Taeniolabis taoensis
Восстановление Taeniolabis taoensis В своем исследовании 2001 г. Kielan-Jaworowska и Hurum обнаружили, что большинство мультитуберкулятов можно отнести к двум подотрядам: « Plagiaulacida » и « Cimolodonta». Исключение составляет род Arginbaatar, который имеет общие характеристики с обеими группами.
«Plagiaulacida» - парафилетическая, представляющая более примитивную эволюционную ступень и, возможно, более производную гондванатерию. Его представители - более базальные Multituberculata, хотя гондванатерии являются скорее производными. Хронологически они варьировались от среднего юрского периода (безымянный материал) до нижнего мела. Эта группа подразделяется на три неформальных группировок: allodontid линии, то paulchoffatiid линии и plagiaulacid линии.
Гондванатерии является монофилетическая группа, которая была разнообразной в позднем мелу в Южной Америке, Индии, Мадагаскара и, возможно, Африке и происходит далее в кайнозое из Южной Америки и Антарктиды. Хотя их принадлежность к мультитуберкулезу оспаривается, последние филогенетические исследования показывают, что они являются сестринской группой цимолодонтов. Есть два основных семейства, Ferugliotheriidae и Sudamericidae, с неопределенным расположением нескольких таксонов, таких как Greniodon и Groeberia. Патагония - это самый молодой известный мультитуберкулез, возникший в миоцене Аргентины.
Cimolodonta, по-видимому, является естественным ( монофилетическим ) подотрядом. Сюда входят более производные Multituberculata, которые были идентифицированы от нижнего мела до эоцена. Выделяются надсемейства Djadochtatherioidea, Taeniolabidoidea, Ptilodontoidea, а также группа Paracimexomys. Кроме того, существуют семейства Cimolomyidae, Boffiidae, Eucosmodontidae, Kogaionidae, Microcosmodontidae и два рода Узбекбаатар и Viridomys. Более точное размещение этих типов требует дальнейших открытий и анализа.
| Подгруппы | ||||
|---|---|---|---|---|
 Многотуберкулезное филогенетическое дерево Основная статья: Список многотуберкулезных видов Многотуберкулезное филогенетическое дерево Основная статья: Список многотуберкулезных видов На основе совместных работ Mikko's Phylogeny Archive и Paleofile.com. Подотряд † Plagiaulacida Simpson 1925
|
| Мультитуберкулята |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Мультитуберкуляты существовали около 166 или 183 миллионов лет и часто считаются самыми успешными, разнообразными и долгоживущими млекопитающими в естественной истории. Впервые они появились в юрский, или, возможно, даже триаса, пережили массовое вымирание в меловом периоде, и вымерли в начале олигоцена эпохи, около 35 миллионов лет назад. Самый старый из известных видов в группе - это Indobaatar zofiae из юрского периода Индии, около 183 миллионов лет назад, а самыми молодыми являются два вида, Ectypodus lovei и безымянный возможный неоплагиулацид, из отложений позднего эоцена / олигоцена Медисин- Пул- Хиллз в Северной Дакоте.. Если гондванатеры являются мультитуберкулезными, то клады, возможно, выжили еще дольше в колхуэуапском миоцене в Южной Америке в форме Patagonia peregrina.
Мультитуберкуляты - одни из первых млекопитающих, демонстрирующих сложное социальное поведение.
Исчезновение мультитуберкулеза было предметом споров в течение нескольких десятилетий. После, по крайней мере, 88 миллионов лет доминирования над большинством сообществ млекопитающих, мультитуберкуляты достигли пика своего разнообразия в раннем палеоцене, прежде чем постепенно уменьшаться на последних этапах эпохи и в эоцене, окончательно исчезнув в раннем олигоцене (в середине миоцена, если гондванатерии - мультитуберкулезные). Традиционно вымирание мультитуберкулятов связывали с ростом грызунов (и, в меньшей степени, более ранних конкурентов плаценты, таких как hyopsodonts и Plesiadapiformes ), которые предположительно исключали мультитуберкулезных организмов из большинства фаун млекопитающих.
Однако идея о том, что мультитуберкуляты были заменены грызунами и другими плацентарями, подвергалась критике со стороны нескольких авторов. Во-первых, он основан на предположении, что эти млекопитающие «уступают» более производным плацентарям, и игнорирует тот факт, что грызуны и мультитуберкуляты сосуществовали вместе по крайней мере 15 миллионов лет. По мнению некоторых исследователей, «упадок» мультитуберкулеза формируется резкими событиями вымирания, особенно после тиффанского периода, когда происходит внезапное падение разнообразия. Наконец, самые молодые известные мультитуберкулезные заболевания не являются примером моделей конкурентного исключения; Олигоценовый Ectypodus - скорее универсальный вид, чем специалист. Эта комбинация факторов предполагает, что вместо постепенного снижения из-за давления со стороны грызунов и аналогичных плацентарных существ, мультитуберкулезные животные просто не могли справиться с климатическими и растительными изменениями, а также с появлением новых хищных эутерианцев, таких как миациды.
Более поздние исследования показывают неоднозначный эффект. Многотуберкулезные фауны в Северной Америке и Европе действительно снижают корреляцию с интродукцией грызунов в эти районы. Однако азиатские многотуберкулезные фауны сосуществовали с грызунами с минимальным количеством случаев вымирания, подразумевая, что конкуренция не была основной причиной исчезновения азиатских мультитуберкулезных животных. В целом кажется, что азиатские мультитуберкуляты, в отличие от североамериканских и европейских видов, так и не оправились от события KT, что в первую очередь позволило эволюционировать и размножаться грызунам. Недавнее исследование, похоже, действительно указывает на то, что эутерианцы быстрее выздоравливали после события КТ, чем мультитуберкулезные. И наоборот, другое исследование показало, что плацентарное облучение не начиналось в значительной степени до тех пор, пока не снизилось количество мульти-туберкулёзов.
Конкуренция между гондванатериями и грызунами и / или другими глирами не проверена, с большим промежутком времени между самыми молодыми представителями первых в Индии, Африке и Мадагаскаре в маастрихте и первыми представителями последних в палеоцене, эоцене и олигоцене соответственно.. Сосуществование между обеими группами в настоящее время подтверждено только в Южной Америке, Патагония Перегрин, как полагает, были вынуждены в специализированной роющую нишу по конкуренции с грызунами и argyrolagoidean paucituberculate сумчатым, а другие клады, Groeberiidae, достиг своего пик разнообразия в середине Олигоцен, после прибытия грызунов.
Млекопитающее Saint Bathans, небольшое ископаемое млекопитающего известно из миоцена (Manuherikia Group, датированный 19 миллионов лет назад) в Новой Зеландии, возможно, было многобугорчатым. Если это так, то это самый последний из существующих мультитуберкулезных заболеваний. Вероятно, это реликтовый вид, который обитал в Новой Зеландии, когда она отделилась от Гондваны 80 миллионов лет назад.
Мультитуберкулезные заболевания в основном известны с северных континентов ( Лавразия ), но есть различные записи с южных континентов ( Гондвана ). Группа Gondwanatheria, известная из Аргентины, Антарктиды, Мадагаскара, Индии и, возможно, Танзании, была отнесена к отряду в прошлом, и, хотя это положение остается спорным, самые последние филогенетические исследования выявили их как мультитуберкулезные за пределами, но близко к Cimolodonta. Два рода, Hahnodon и Denisodon, известны из раннего мела Марокко, но вместо этого они могут быть харамииданами. Мультитуберкулезные заболевания также были зарегистрированы в позднем меловом периоде Мадагаскара и Аргентины. Австралийский мульти-туберкулез, Corriebaatar, известен по единственному зубу. Индобаатар известен из формации Кота в Индии, которая тогда входила в состав восточной части Гондваны, и является старейшим известным многотуберкулезным заболеванием.
В позднем меловом периоде мультитуберкуляты были широко распространены и разнообразны в северном полушарии и, возможно, также на большинстве южных массивов суши, составляя более половины видов млекопитающих типичной фауны. Хотя несколько линий вымерли во время круговорота фауны в конце мелового периода, мультитуберкулятам в целом удалось очень успешно пересечь границу мела и палеогена и достичь пика своего разнообразия в палеоцене. Они были важным компонентом почти всех палеоценовых фаун Европы и Северной Америки, а также некоторых позднепалеоценовых фаун Азии. Мультитуберкуляты также были самыми разнообразными по размеру в палеоцене - от размера очень маленькой мыши до размера панды. Однако в Азии мультитуберкуляты палеоцена и эоцена составляют очень небольшой процент от общей местной фауны млекопитающих, им никогда не удавалось оправиться от события KT так, как это сделали их североамериканские и европейские аналоги. Gondwanatheres распространены в позднем мелу из Мадагаскара и Индии, в палеоцена и эоцена на острове Сеймур, и происходят в Южной Америке от позднего мела до миоцена.