
Войти
 1: более мелкие (желтые) виды птиц кормятся по всему дереву.. 2: более крупные (красные) виды конкурируют за ресурсы.. 3: красный цвет преобладает в средний для более богатых ресурсов. Желтый адаптируется к новой нише, ограниченной вершиной и основанием дерева, избегая конкуренции.
1: более мелкие (желтые) виды птиц кормятся по всему дереву.. 2: более крупные (красные) виды конкурируют за ресурсы.. 3: красный цвет преобладает в средний для более богатых ресурсов. Желтый адаптируется к новой нише, ограниченной вершиной и основанием дерева, избегая конкуренции.В экологии используется принцип конкурентного исключения, иногда обозначаемый как Закон Гаузе - это предложение, названное в честь Георгия Гаузе, что два вида , конкурирующие за один и тот же ограниченный ресурс, не могут сосуществовать в постоянные значения популяции. Когда один вид имеет хоть малейшее преимущество перед другим, в долгосрочной перспективе доминирует тот, который имеет это преимущество. Это ведет либо к исчезновению более слабого конкурента, либо к эволюционному или поведенческому сдвигу в сторону другой экологической ниши. Принцип был перефразирован в изречении «полные конкуренты не могут сосуществовать».
Принцип конкурентного исключения классически приписывается Георгию Гаузе, хотя он фактически никогда не сформулировал его. Этот принцип уже присутствует в теории естественного отбора Дарвина.
На протяжении всей своей истории статус принципа колебался между априори («два сосуществующих вида должны иметь разные ниши») и экспериментальной истиной («мы находим эти сосуществующие виды имеют разные ниши »).
 Paramecium aurelia и Paramecium caudatum хорошо растут по отдельности, но когда они конкурируют за одни и те же ресурсы, P. aurelia превосходит P. caudatum.
Paramecium aurelia и Paramecium caudatum хорошо растут по отдельности, но когда они конкурируют за одни и те же ресурсы, P. aurelia превосходит P. caudatum. Основываясь на полевых наблюдениях, Джозеф Гриннелл сформулировал принцип конкурентного исключения в 1904 году: «Маловероятно, что два вида с примерно одинаковыми пищевыми привычками будут долго оставаться равномерно сбалансированными по численности. в одном регионе. Одно будет вытеснять другое ". Георгий Гаузе сформулировал закон конкурентного исключения на основе лабораторных экспериментов по конкуренции с использованием двух видов Paramecium, P. aurelia и P. caudatum. Условия заключались в том, чтобы добавлять свежую воду каждый день и вводить постоянный поток пищи. Хотя первоначально доминировал P. caudatum, P. aurelia восстановилась и впоследствии способствовала вымиранию P. caudatum посредством эксплуататорской ресурсной конкуренции. Однако Гаузе удалось позволить P. caudatum выжить за счет различных параметров окружающей среды (еда, вода). Таким образом, закон Гаузе справедлив, только если экологические факторы постоянны.
Гаузе также изучал конкуренцию между двумя видами дрожжей, обнаружив, что Saccharomyces cerevisiae постоянно побеждает, производя более высокую концентрацию этилового спирта.
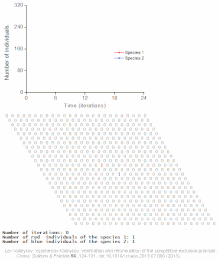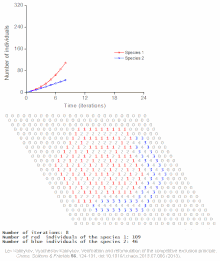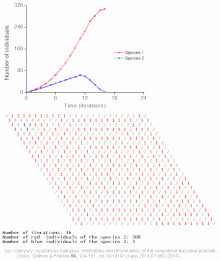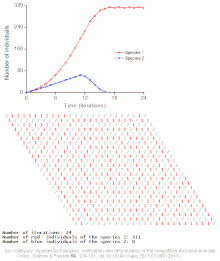 Клеточный автомат модель межвидовой конкуренции за один ограниченный ресурс
Клеточный автомат модель межвидовой конкуренции за один ограниченный ресурс Конкурентное исключение предсказывается математическими и теоретическими моделями, такими как модели Лотки-Вольтерры конкуренции. Однако по малоизученным причинам конкурентное исключение редко наблюдается в природных экосистемах, и многие биологические сообщества, похоже, нарушают закон Гаузе. Самый известный пример - так называемый «парадокс планктона ». Все виды планктона живут за счет очень ограниченного количества ресурсов, в первую очередь солнечной энергии и минералов, растворенных в воде. Согласно принципу конкурентного исключения, только небольшое количество видов планктона должно иметь возможность сосуществовать на этих ресурсах. Тем не менее, большое количество видов планктона сосуществует в небольших районах открытого моря.
Некоторыми общинами, которые, по-видимому, придерживаются принципа конкурентного исключения, являются певчие птицы Макартура певчие и зяблики Дарвина, хотя последние по-прежнему очень сильно пересекаются с экологической точки зрения. сильно, поскольку конкуренция отрицательно влияет только на них в экстремальных условиях.
Частичное решение парадокса заключается в повышении размерности системы. Пространственная неоднородность, трофические взаимодействия, множественная ресурсная конкуренция, компромисс между конкуренцией и колонизацией и отставание могут предотвратить исключение (игнорирование стохастического вымирания на более длительных таймфреймах). Однако такие системы, как правило, трудноразрешимы с аналитической точки зрения. Кроме того, многие теоретически могут поддерживать неограниченное количество видов. Возникает новый парадокс: большинство известных моделей, допускающих стабильное сосуществование, допускают сосуществование неограниченного числа видов, однако в природе любое сообщество содержит лишь горстку видов.
Недавние исследования, посвященные некоторым предположениям, сделанным для моделей, предсказывающих конкурентное исключение, показали, что эти предположения необходимо пересмотреть. Например, небольшая модификация предположения о том, как связаны рост и размер тела, приводит к другому выводу, а именно к тому, что для данной экосистемы определенный диапазон видов может сосуществовать, в то время как другие становятся вытесненными.
Один Одним из основных способов сосуществования видов, разделяющих нишу, является компромисс между конкуренцией и колонизацией. Другими словами, виды, которые являются лучшими конкурентами, будут специалистами, тогда как виды, которые являются лучшими колонизаторами, с большей вероятностью будут универсальными. Модели «хост-паразит» - эффективные способы изучения этой взаимосвязи с использованием событий передачи хоста. Кажется, есть два места, где способность к колонизации у экологически близких видов различается. Что касается перьевых вшей, Буш и Клейтон представили некоторую проверку этого, показав, что два близкородственных рода вшей почти равны по своей способности колонизировать новых голубей-хозяев после переноса. Харбисон продолжил эту линию мысли, исследуя, различаются ли два рода по способности к передаче. Это исследование было сосредоточено в первую очередь на определении того, как происходит колонизация и почему крылатые вши лучше колонизируют, чем тельца. Вертикальный перенос является наиболее частым явлением между родителем и потомством, он хорошо изучен и хорошо изучен. Горизонтальный перенос трудно измерить, но у вшей, похоже, происходит форез или «автостоп» одного вида за другим. Харбисон обнаружил, что тельца менее искусны в форезе и лучше других конкурируют, тогда как крылатые вши преуспевают в колонизации.
экологическое сообщество - это совокупность видов, которая поддерживается экологическим (Hutchinson, 1959; Leibold, 1988) и эволюционным процессом (Weiher and Keddy, 1995; Chase et al., 2003). Эти два процесса играют важную роль в формировании существующего сообщества и будут продолжаться в будущем (Tofts et al., 2000; Ackerly, 2003; Reich et al., 2003). В местном сообществе потенциальные члены фильтруются сначала по факторам окружающей среды, таким как температура или наличие необходимых ресурсов, а затем, во-вторых, по его способности сосуществовать с другими резидентами.
В подходе к пониманию того, как два вида сочетаются друг с другом в сообществе или как все сообщество объединяется, «Происхождение видов» (Дарвин, 1859 г.) предполагает, что при однородных условиях окружающей среды борьба за Существование больше между близкородственными видами, чем между отдаленно родственными видами. Он также предположил, что функциональные черты могут сохраняться при разных филогенезах. Такое сильное филогенетическое сходство между близкородственными видами известно как филогенетические эффекты (Derrickson et al., 1988.)
С помощью полевых исследований и математических моделей эколог установил связь между сходством функциональных признаков между видами и его влиянием. о сосуществовании видов. Согласно гипотезе конкурентного родства (Cahil et al., 2008) или филогенетической гипотезе ограничивающего сходства (Violle et al., 2011) межвидовая конкуренция высока среди видов, которые имеют сходные функциональные признаки и которые конкурируют за аналогичные ресурсы и среду обитания. Следовательно, это вызывает сокращение числа близкородственных видов и даже их распространение, известное как филогенетическая сверхдисперсия (Webb et al., 2002). Обратной стороной филогенетической избыточной дисперсии является филогенетическая кластеризация, в этом случае ожидается, что виды с консервативными функциональными признаками будут совместно встречаться из-за фильтрации окружающей среды (Weiher et al., 1 995; Webb, 2000). В исследовании, проведенном Webb et al., 2000, они показали, что небольшие участки леса Борнео содержат вместе тесно связанные деревья. Это говорит о том, что у близкородственных видов общие черты, которым благоприятствуют конкретные факторы окружающей среды, которые различаются между участками, вызывающими филогенетическую кластеризацию.
Для обоих филогенетических паттернов (филогенетическая сверхдисперсия и филогенетическая кластеризация) исходное предположение состоит в том, что филогенетически родственные виды также имеют экологическое сходство (H. Burns et al, 2011). Нет значительного количества экспериментов, позволяющих определить, насколько близкородственные виды сходны по нише. В связи с этим интерпретировать оба филогенетических паттерна непросто. Было показано, что филогенетическая чрезмерная дисперсия может также быть результатом конвергенции отдаленных родственных видов (Cavender-Bares et al. 2004; Kraft et al. 2007). В своем исследовании они показали, что черты сходятся, а не сохраняются. В то время как в другом исследовании было показано, что филогенетическая кластеризация также может быть связана с историческими или биогеографическими факторами, которые не позволяют видам покинуть свои предковые ареалы. Таким образом, необходимы дополнительные филогенетические эксперименты для понимания силы взаимодействия видов в сообществе.
Доказательства того, что принцип конкурентного исключения действует в человеческих группах, были проанализированы и интегрированы в теорию реальности для объяснения воинственных и мирных обществ. Например, группы охотников-собирателей, окруженные другими группами охотников-собирателей в той же экологической нише, будут сражаться, по крайней мере, время от времени, в то время как группы охотников-собирателей, окруженные группами с другими средствами к существованию, могут мирно сосуществовать.