
Войти
| Heterodontosauridae. Временной диапазон: Ранняя юра - Ранний мел, 200–133 Ma До Ꞓ O S D C P T J K Pg N Возможная Поздняя триасовая запись | |
|---|---|
 | |
| Отливка образца SAM-PK-K1332 из Heterodontosaurus tucki, Музей палеонтологии Калифорнийского университета | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Chordata |
| Clade: | Dinosauria |
| Отряд: | †Ornithischia |
| Семейство: | †Heterodontosauridae. Romer, 1966 |
| Подгруппы | |
| Синонимы | |
| |
Heterodontosauridae - это семейство ранних орнитисхийских динозавров, которые, вероятно, были одними из самых базальных (примитивных) Члены группы. Хотя их окаменелости относительно редки, а их группа небольшая по численности, они жили на всех континентах, кроме Австралии, примерно 100 миллионов лет, от позднего триаса до Раннемеловые.
Гетеродонтозавриды были лисицами динозаврами длиной менее 2 метров (6,6 футов), включая длинный хвост. Они известны в основном своими характерными зубами, в том числе увеличенными клыками -подобными бивнями и щечными зубами, приспособленными для жевания, аналогичными зубам меловых гадрозаврид. Их рацион был травоядным или, возможно, всеядным.
 Сравнение размеров многих гетеродонтозаврид
Сравнение размеров многих гетеродонтозаврид  Восстановление жизни Fruitadens
Восстановление жизни Fruitadens Среди гетеродонтозаврид только сам гетеродонтозавр известен по полному скелету. Фрагментарные останки скелета Abrictosaurus известны, но не были полностью описаны, в то время как большинство других гетеродонтозаврид известны только по фрагментам челюстей и зубов. Следовательно, большинство гетеродонтозаврид синапоморфий (определяющих признаков) было описано на основе зубов и костей челюсти. Гетеродонтозавр имел длину чуть более 1 метра (3,3 фута), в то время как фрагментарные останки Lycorhinus могут указывать на более крупную особь.
Tianyulong из Китая, по-видимому, сохраняет нитчатые покровы, которые, как считается, быть вариантом прото-перьев, обнаруженных у некоторых теропод. Эти волокна включают гребень вдоль хвоста. Наличие этих нитчатых покровов позволяет предположить, что и орнитисхи, и заурисхии были эндотермическими.
У абриктозавра и гетеродонтозавра были очень большие глаза. Под глазами скуловая кость выступала вбок, эта особенность также присутствует у цератопсов. Как и в челюстях большинства орнитисхийцев, передний край предчелюстной кости (кость на кончике верхней челюсти) был беззубым и, вероятно, поддерживал ороговевший клюв (rhamphotheca ), хотя у гетеродонтозаврид действительно были зубы в заднем отделе предчелюстной кости. Большой промежуток, называемый диастемой, отделял эти предчелюстные зубы от зубов верхней челюсти (основная кость верхней челюсти) у многих орнитисхов, но эта диастема была характерна для гетеродонтозаврид. нижняя челюсть (нижняя челюсть) была закруглена predentary, костью, уникальной для орнитизхий. Эта кость также поддерживала клюв, подобный тому, который находится на предчелюстной кости. Все зубы нижней челюсти были обнаружены на зубной кости.
гетеродонтозаврид названы в честь их сильно гетеродонта зубного ряда. Три предчелюстных зуба. В ранней юре Abrictosaurus, Heterodontosaurus и Lycorhinus первые два предчелюстных зуба были маленькими и коническими, в то время как гораздо более крупный третий зуб напоминал клыки карниворанов млекопитающих и часто называют клыковидной формой или «бивнем». Нижний клыковидный, крупнее верхнего, занял первое положение в зубной кости и был размещен арочной диастемой верхней челюсти при закрытии рта. Эти caniniforms имели зубцы как на переднем, так и на заднем крае у Heterodontosaurus и Lycorhinus, в то время как у Abrictosaurus зазубрины были только на переднем крае. В раннемеловом Echinodon, возможно, было два верхних клыковидных, которые были на верхней челюсти, а не на предчелюстной кости, а у Fruitadens из поздней юры могло быть по две нижних клыковидной формы на каждой. dentary.
 Эволюция ключевых жевательных специализаций гетеродонтозаврид, согласно Sereno, 2012
Эволюция ключевых жевательных специализаций гетеродонтозаврид, согласно Sereno, 2012 Подобно характерным бивням, щечные зубы производных гетеродонтозаврид также были уникальными среди ранних орнитисхов. Небольшие гребни, или зубчики, выстилали края орнитисхиевых щечных зубов, чтобы обрезать растительность. У всех гетеродонтозаврид эти зубчики проходят только на треть вниз по коронке зуба от кончика; у других орнитисхий зубчики проходят дальше вниз к корню. Базальные формы, такие как абриктозавр, имели щечные зубы на верхней и нижней челюсти, которые в целом были похожи на других орнитисхий: широко расставленные, у каждой была низкая коронка и сильно развитый гребень (поясная корка), отделяющий коронку от корня. У более производных форм, таких как Lycorhinus и Heterodontosaurus, зубы имели форму долота, с гораздо более высокими коронками и без цингулы, так что не было разницы в ширине между коронками и корнями.
Эти производные щечные зубы накладывались друг на друга, так что их коронки образовывали непрерывную поверхность, на которой можно было жевать пищу. Зубные ряды были слегка вставлены со стороны рта, оставляя за пределами зубов пространство, которое могло быть ограничено мышечной щекой, которая была необходима для жевания. Гадрозавры и цератопсы мелового периода, а также многие травоядные млекопитающие одновременно эволюционируют в некоторой степени аналогичные зубные батареи. В отличие от гадрозавров, у которых постоянно заменялись сотни зубов, замена зубов у гетеродонтозаврид происходила гораздо медленнее, и несколько экземпляров были обнаружены без единого ожидающего замены зуба. Характерно, что гетеродонтозавриды не имели небольших отверстий (отверстий) на внутренней стороне костей челюсти, которые, как полагают, способствовали развитию зубов у большинства других орнитисхий. Гетеродонтозавриды также могли похвастаться уникальным сфероидальным соединением между зубными и предзубными зубами, что позволяло нижним челюстям вращаться наружу при закрытии рта, прижимаясь щеками друг к другу. Из-за низкой скорости замены такое шлифование приводило к сильному износу зубов, который обычно стирал большую часть зубцов на старых зубах, хотя увеличенная высота коронок обеспечила каждому зубу долгую жизнь.
 Ископаемое Тяньюлонг, морда, рука, ступни и хвост в красной рамке
Ископаемое Тяньюлонг, морда, рука, ступни и хвост в красной рамке Посткраниальная анатомия Heterodontosaurus tucki хорошо описана, хотя H. tucki обычно считается наиболее производным от раннеюрских гетеродонтозаврид, поэтому невозможно узнать, как многие из его черт были общими с другими видами. У динозавра передние конечности были длинными, более 70% от длины задних. Хорошо развитый дельтовидный гребень (гребень для прикрепления грудных и плечевых мышц) плечевой кости и выступающего локтевого отростка ( где были прикреплены мышцы, расширяющие предплечье ) локтевой кости, что указывает на то, что передняя конечность также была сильной. На manus («рука») было пять цифр. Первый был большим, с остро изогнутым когтем и вращался внутрь при сгибании; Роберт Баккер назвал это «поворотным пальцем». Вторая цифра была самой длинной, немного длиннее третьей. Оба эти пальца были с когтями, в то время как четвертый и пятый пальцы были очень маленькими и простыми по сравнению. В задней конечности большеберцовая кость была на 30% длиннее, чем бедренная кость, что обычно считается приспособлением для скорости. Большеберцовая кость и малоберцовая кость голени слились с астрагалом и пяточной костью голеностопного сустава, образуя «тибиофибиотарсус » конвергентно с современные птицы. Также, как и у птиц, нижние предплюсневые кости (лодыжки) и плюсневые кости были слиты с образованием «tarsometatarsus ». В pes (задняя лапа) четыре цифры, при этом только вторая, третья и четвертая касаются земли. Хвост, в отличие от многих других орнитисхийцев, не имел окостенелых сухожилий для сохранения жесткой позы и, вероятно, был гибким. Фрагментарный скелет, известный как Abrictosaurus, никогда не был полностью описан, хотя передняя конечность и кисть были меньше, чем у Heterodontosaurus. Кроме того, каждый четвертый и пятый пальцы передней конечности имеют на одну фаланговую кость.
 Голотип челюсть геранозавра
Голотип челюсть геранозавра южноафриканского палеонтолога Роберт Брум создал название Geranosaurus в 1911 году для костей челюсти динозавров, у которых отсутствуют все зубы и некоторые частично связанные кости конечностей. В 1924 году Lycorhinus был назван и классифицирован как цинодонт Сидни Хотоном. Гетеродонтозавр был назван в 1962 году, и его, Lycorhinus и Geranosaurus были признаны близкородственными орнитисхийскими динозаврами. Альфред Ромер назвал Heterodontosauridae в 1966 году семейством орнитисхийских динозавров, включая Heterodontosaurus и Lycorhinus. Он был определен как клады в 1998 году Полом Серено и переопределен им в 2005 году как стволовая клада, состоящая из Heterodontosaurus tucki и всех видов, более близких к Heterodontosaurus, чем к Parasaurolophus walkeri, Pachycephalosaurus wyomingensis, Triceratops horridus или Ankylosaurus magniventris.
Heterodontosauridae включает роды Abrictosaurus, Lycorhinus и Heterodonon из Южной Африки. В то время как Ричард Талборн однажды переназначил всех троих Ликорину, все остальные авторы считают эти три рода разными. Внутри семейства Heterodontosaurus и Lycorhinus считаются сестринскими таксонами, а абриктозавр является базальным членом. Геранозавр также является гетеродонтозавридом, но обычно считается nomen dubium, потому что у типового экземпляра отсутствуют все зубы, что делает его неотличимым от любого другого рода в семействе. Совсем недавно род Echinodon был признан гетеродонтозавридом в нескольких исследованиях. Lanasaurus был назван в честь верхней челюсти в 1975 году, но более поздние открытия показали, что он принадлежит Lycorhinus, что сделало Lanasaurus младший синоним этого рода. Dianchungosaurus когда-то считался гетеродонтозавридом из Азии, но с тех пор было показано, что останки были химерой из прозауропод и мезоэвкрокодилий остались. Хосе Бонапарт также однажды классифицировал южноамериканского пизанозавра как гетеродонтозаврида, но теперь это животное известно
 Череп Abrictosaurus
Череп Abrictosaurus Принадлежность к Heterodontosauridae хорошо установлена по сравнению с его неопределенным филогенетическим положением. Несколько ранних исследований показали, что гетеродонтозавриды были очень примитивными орнитишами. Из-за предполагаемого сходства морфологии передних конечностей Роберт Баккер предложил связь между гетеродонтозавридами и ранними зауроподоморфами, такими как Anchisaurus, соединяя отряды Saurischia и Ornithischia. Доминирующая гипотеза последних нескольких десятилетий относила гетеродонтозаврид к базальным орнитоподам. Однако другие предположили, что гетеродонтозавриды имеют общего предка с Marginocephalia (цератопсы и пахицефалозавры ), и эта гипотеза нашла поддержку в некоторых исследованиях начала 21 века. Клада, содержащая гетеродонтозаврид и маркоцефалов, получила название Heterodontosauriformes. Гетеродонтозавриды также рассматривались как базальные как для орнитопод, так и для маргоцефалов. В 2007 году кладистический анализ показал, что гетеродонтозавриды являются базальными по отношению ко всем известным орнитишиям, кроме пизанозавра, что перекликается с некоторыми из самых ранних работ по семейству. Однако исследование Бонапарта показало, что Pisanosauridae являются синонимом Heterodontosauridae, а не отдельным семейством, в результате чего Pisanosaurus является гетеродонтозавром. Батлер и др. (2010) обнаружили, что Heterodontosauridae являются наиболее распространенным из известных орнитисхиальных излучений.
кладограмма ниже показывает взаимосвязь внутри Heterodontosauridae и следует за анализом Sereno, 2012:
| Heterodontosauridae |
| |||||||||||||||||||||||||||||||||||||||

 Биогеографическое распределение гетеродонтозаврид во времени
Биогеографическое распределение гетеродонтозаврид во времени Хотя первоначально известно остатки гетеродонтозаврид из ранней юры на юге Африки теперь известны с четырех континентов. В начале истории гетеродонтозаврид суперконтинент Пангея все еще был в значительной степени нетронутым, что позволило семье достичь почти всемирного распространения. Самые старые из известных возможных останков гетеродонтозаврид - это фрагмент челюсти и отдельные зубы из формации Лагуна Колорада в Аргентине, которая восходит к позднему триасу. Эти останки имеют производную морфологию, аналогичную гетеродонтозавру, включая собачковидную форму с зубцами как на переднем, так и на заднем крае, а также высокие корончатые зубы верхней челюсти без цингулюма. Ирмис и др. (2007) предварительно согласились с тем, что этот ископаемый материал представляет собой гетеродонтозаврид, но заявили, что для подтверждения этого предположения необходим дополнительный материал, потому что образец плохо сохранился, в то время как Серено (2012) только заявил, что этот материал может представлять орнитисхий или даже гетеродонтозаврид. Olsen, Kent Whiteside (2010) отметили, что возраст самой формации Laguna Colorada плохо ограничен, и поэтому не было окончательно определено, имеет ли предполагаемый гетеродонтозаврид из этой формации триасовый или юрский возраст.. Самая разнообразная гетеродонтозаврид фауна происходит из ранней юры на юге Африки, где обнаружены окаменелости гетеродонтозавра, абриктозавра, ликоринуса и сомнительного геранозавра.
Неописанные раннеюрские гетеродонтозавры также известны по США и Мексика соответственно. Кроме того, начиная с 1970-х годов, большое количество окаменелостей было обнаружено в позднеюрской формации Моррисон около Фруита, Колорадо в Соединенных Штатах. Описанный в печати в 2009 году, этот материал был отнесен к роду Fruitadens. Зубы гетеродонтозаврид, лишенные цингулюма, также были описаны в позднеюрских и раннемеловых образованиях Испании и Португалии. Останки эхинодона были переописаны в 2002 году, что показало, что он может представлять позднего выжившего гетеродонтозаврида из берриаса стадии раннего мела на юге Англии. Dianchungosaurus из ранней юры Китая больше не считается гетеродонтозавридом; хотя известна одна сохранившаяся поздно азиатская форма (Тианюлонг ).
 Восстановление Пегомастакс
Восстановление Пегомастакс Большинство окаменелостей гетеродонтозаврид обнаружено в геологических формациях, которые представляют собой засушливые или полузасушливые среды, в том числе формация Верхний Эллиот в Южной Африке и слои Пурбек на юге Англии. Было высказано предположение, что гетеродонтозаврид подвергался сезонной лету или спячка в самое засушливое время года. Из-за отсутствия замещающих зубов у большинства гетеродонтозаврид было предложено заменить весь набор зубов в течение этого периода покоя, поскольку это казалось непрерывным и спорадическим Замена зубов прервала бы функцию зубного ряда как единой жевательной поверхности. Однако это было основано на неправильном понимании механики челюстей гетеродонтозаврид. Считалось, что гетеродонтозавриды действительно заменяли свои зубы постоянно, хотя и медленнее, чем у других рептилии, но КТ черепов молодых и зрелых гетеродонтозавров не обнаруживает замещающих зубов. В настоящее время нет доказательств, подтверждающих гипотезу об летеврении у гетеродонтозаврид, но она не может быть отвергнута на основании сканированных изображений черепа.
В то время как щечные зубы гетеродонтозаврид явно приспособлены для жесткого скрежета. растительный материал, их диета могла быть всеядной. Заостренные передчелюстные зубы и острые изогнутые когти на передних конечностях предполагают некоторую степень хищничества. Было высказано предположение, что длинные мощные передние конечности гетеродонтозавра могли быть полезны для разрыва насекомых в гнездах, подобно современным муравьедам. Эти передние конечности также могли использоваться в качестве инструментов для выкапывания, возможно, для корней и клубней.
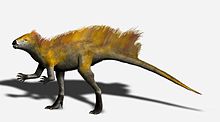 восстановления тяньюлонга
восстановления тяньюлонга Длина передней конечности по сравнению с задней конечностью предполагает, что гетеродонтозавр мог быть частично Четвероногие, а выдающийся олекранонный отросток и сверхразгибаемые пальцы передних конечностей встречаются у многих четвероногих. Однако рука явно предназначена для захвата, а не для поддержки веса. Многие особенности задней конечности, в том числе длинная большеберцовая кость и ступня, а также слияние тибиофибиотарсуса и тарсометатарсуса, указывают на то, что гетеродонтозавры были приспособлены к быстрому бегу на задних конечностях, поэтому маловероятно, что гетеродонтозавр передвигался на всех четырех конечностях, за исключением, возможно, случая
Короткие бивни, найденные у всех известных гетеродонтозаврид, сильно напоминают бивни, найденные у современных кабарги, пекари и свиней. У многих из этих животных (а также у длинноберых моржей и азиатских слонов ) это половой диморфный признак, причем бивни встречаются только у самцов.. У экземпляра типа абриктозавра отсутствуют бивни, и первоначально он был описан как женщина. Хотя это остается возможным, несросшиеся крестцовые позвонки и короткая морда указывают на то, что этот экземпляр представляет собой молодое животное. Второй, более крупный экземпляр, первоначально предположительно принадлежавший к абриктозавру, явно обладает бивнями, что было использовано для подтверждения идеи о том, что бивни встречаются только у взрослых, а не являются вторичной сексуальной характеристикой самцов. Эти клыки могли быть использованы для боя или показа с представителями того же вида или с другими видами. Отсутствие бивней у молодых абриктозавров также может быть еще одной характеристикой, отделяющей его от других гетеродонтозавров, поскольку бивни известны у молодых гетеродонтозавров. Другие предлагаемые функции бивней включают защиту и использование в иногда всеядной диете. Однако в 2012 году Серено в качестве альтернативы назначил этот экземпляр Lycorhinus, который, как уже известно, обладал бивнями, и поэтому их отсутствие у абриктозавра могло не быть результатом возраста.
В 2005 г. В Южной Африке был обнаружен небольшой полный окаменевший скелет гетеродонтозаврид возрастом более 200 миллионов лет. В июле 2016 года он был сканирован группой исследователей из Южной Африки с использованием Европейского центра синхротронного излучения ; сканирование зубных рядов выявило кости неба толщиной менее миллиметра.