| Гудзонелевпидия <779ue>Еврологи 807>Ихтиозаврота <779ue>Еврологи 807>Ихтиозаврота ОписаниеРазмерИхтиозавров в среднем около 6–4 м (в среднем около 6–4 м) –13,1 фута) в длину. Отдельные экземпляры были всего 0,3 м (1 фут); некоторые виды были намного крупнее: триасовый Shonisaurus popularis был около 15 м (49 футов) в длину, а в 2004 г. Shastasaurus sikanniensis (иногда классифицируемый как Shonisaurus sikanniensis), по оценкам, составлял 21 м (69 футов) в длину. Фрагментарные находки предполагают формы длиной 15 метров (49 футов) в ранней юре. В 2018 году сообщалось, что фрагменты нижней челюсти из Англии имели длину от 20 до 25 м (от 66 до 82 футов). Согласно оценкам, Stenopterygius длиной 2,4 м (8 футов) весил около 163–168 кг (359–370 фунтов), в то время как 4-метровый весый (13 футов) Ophthalmosaurus icenicus весил 930–950 кг (2 050–370 фунтов). Общая конструкция CGI-реставрация Ichthyosaurus communis CGI-реставрация Ichthyosaurus communis В то время как самые ранние известные представители линии ихтиозавров были похожи на угря по телосложению, более поздние ихтиозавры напоминают более типичных рыб или дельфины, имеющую голову, похожую на морскую свинью, с короткой шеей и длинной мордой. Передние и задние конечности ихтиозавровровров превратились в полностью превратились в ласты. У некоторых видов был плавник на спине и более или менее вертикальный плавник в задней части довольно короткого хвоста. Хотя ихтиозавры выглядели как рыбы, это не так.  Примеры отличительных черт, присущих как дельфинам, так и производным ихтиоптеригам. Примеры отличительных черт, присущих как дельфинам, так и производным ихтиоптеригам. Эволюционный биолог Стивен Джей Гулд сказал, что ихтиозавр был его любимым примером конвергентной эволюции. где сходство строения аналогично, а не гомологично, следовательно, вызвано не общим происхождением, аналогичной адаптацией к идентичной среде:
"Эта морская рептилия с земными предками так сильно сходились на рыбах Структура тем более примечательными, что они произошли из ничего - у земных рептилий предков не было горба его спина или лезвие на его хвосте, чтобы служить предшественником » Диагностические признакиПроизводные ихтиозавры в узком смысле, как определено Мотани в 1999 году, отличаются от ближайших к ним базальных ихтиоптеригийцев повторно родственниками в определенных чертах. Верхний край глазницы состоит из костного стержня, образованного префронтальной и постфронтальной костями. Боковое крыло крыловидного отростка окостенено не полностью и в разной степени. ала. Задние спинные позвонки имеют дискообразную форму. СкелетЧереп Череп Temnodontosaurus platyodon имеет типичную ихтиозавровую форму с удлиненной мордой и большими глазницами Череп Temnodontosaurus platyodon имеет типичную ихтиозавровую форму с удлиненной мордой и большими глазницами Базальная ихтиоптеригия уже имела удлиненные треугольные черепа. У ихтиозавров в узком смысле их морды стали очень заостренными. Морда образована предчелюстной костью. верхнечелюстная кость позади нее обычно короче и иногда исключена из внешней ноздри вет задней вью предчелюстной кости. Соответственно, количество предчелюстных зубов велико, а верхних зубов меньше или даже полностью отсутствует. Задняя часть морды образована носовыми костями. Производные виды имеют внутреннее отверстие - отверстие по средней линии, разделяющее заднюю часть носовых костей. Носовая кость обычно образует верхний и передний край костной ноздри, которая часто прямо перед глазницей. Однако у некоторых триасовых видов предчелюстная кость настолько сильно вытянута сзади, что даже исключает нос от ноздри.
В задней части черепа преобладает большая глазная впадина, часто закрывающие большую часть задней боковой поверхности. В лунке большое склеральное кольцо ; это круглая структура из небольших перекрывающихся сегментов кости, защищающих глаз от давления воды. И в относительном, и в абсолютном смысле у ихтиозавров самые большие глазницы среди всех известных позвоночных. Другие задние элементы черепа обычно сжаты и слиты, что их идентифицировать. Обычно предполагалось, что верхний задний элемент черепа - это, в то время как чешуйчатая кость и квадратично-скуловая кость иногда сливались. В 1968 году Альфред Шервуд Ромер заявленная предполагаемая надвисочная кость на самом деле чешуйчатой, что было подтверждено в 1973 году Макгоуэном. Однако в 1990 году большинство исследователей убедили, что первоначальная идентификация все-таки была правильной. Надвисочная часть образует задний край надвисочного отверстия; нижнее височное отверстие сбоку отсутствует. Передний край надвисочного отверстия обычно образован постфронтальной частью ; только у самого базального Утацусавр заглазничный и чешуйчатая кость все еще достигают края. Между парными надвисочными отверстиями крыша черепа узкая; у некоторых есть продольный гребень как прикрепление мускулов челюсти. Базальная ихтиоптеригия имеет отверстие теменного глаза между парными теменными костями. У собственно ихтиозавров это перемещается вперед, сначала к границам теменными и лобными видами, в конечном итоге между лобными костями, состояние, показанное производными видами. Загрудинные и пластинчатые кости отсутствуют. Часто кости задней части черепа и неба не полностью окостенели, по-видимому, частично сохранился хрящ. затылочный мыщелок обычно очень выпуклый. стремечко, кость, передающая звуковые волны от барабанной перепонки к среднему уху, удлиненную форму и не проходит через отверстие. Крыловидные зубы обычно отсутствуют. Нижняя челюстьКак и морда, нижняя челюсть удлиненная. У некоторых видов, таких как Eurhinosaurus и Excalibosaurus, передняя часть морды далеко выступает за нижнюю челюсть. В то время как передняя часть нижней челюсти обычно низкая, глубина ее задней части очень изменчива. Большую часть нижней челюсти составляет передняя зубная часть, несущая зуб кость. На своей внутренней стороне зубная кость покрыта селезенкой, которая простирается вперед до симфиза, общей контактной поверхности, на которой срастаются обе нижние челюсти. Суставы челюстей не допускают горизонтальных жевательных движений: они функционируют как простые шарниры, открывающие или закрывающие челюсти по вертикали. ЗубыЗубы ихтиозавров обычно конические. У рыбоядных особей длинные и тонкие коронки зубов слегка загнуты назад. Формы, специализирующиеся на ловле более крупной добычи, имеют более короткие, широкие и прямые зубы; иногда присутствуют режущие кромки. Талаттоархон, высший хищник, имел более крупные зубы, похожие на плоские лезвия. Дурофаги виды, поедавшие моллюсков, имеют низкие выпуклые зубы, которые плотно прилегают друг к другу. Многие зубные зубы ихтиозавров гетеродонты, сочетающие несколько форм зубов, например ряды мелкие зубы спереди и большие зубы сзади. Зубы обычно помещаются в лунки ; производные виды обладают общей зубной бороздой. В последнем случае взрослые люди становятся беззубыми. Зубы в лунках зубов иногда срастаются с челюстной костью. В зубах ихтиозавра дентин видны заметные вертикальные морщины. Дурофаги имеют зубы с глубокими вертикальными бороздками и морщинами в эмали. PostcraniaПозвоночный столб Позвонок ихтиозавра из формыции Sundance (юрский период ) из округа Натрона, штат Вайоминг : Обратите внимание на характерное поперечное сечение в виде песочных часов. (Масштаб в мм.) Позвонок ихтиозавра из формыции Sundance (юрский период ) из округа Натрона, штат Вайоминг : Обратите внимание на характерное поперечное сечение в виде песочных часов. (Масштаб в мм.) Базальная ихтиоптеригия, как и их наземные предки, все еще позвонки, которые обладали полным набором отростков, которые позволяют им сцепляться и сочленяться, образуя позвоночный столб поддерживая вес тела. Оказываемая водой; оказываемая вода; оказываемая вода; другими словами, они были плавучими. Следовательно, позвоночные отростки утратили большую часть своей функции. Собственно ранние ихтиозавры имели задние спинные позвонки, которые стали дискообразными, как у типичных рыб. У более производных видов передние спинные кости также стали дисками. Постепенно утрачивалась большая часть процессов, в том числе прикрепления ребер. Тела позвонков стали намного короче. Передняя и задняя стороны дисков были выдолблены, что произошло к так называемому состоянию. Поперечное сечение такого позвонка имеет форму песочных часов. Эта морфология уникальна для Amniota и позволяет легко отличить позвонки ихтиозавра от позвонков других морских рептилий. Единственный отросток, который сохранил свою функцию, - это позвоночник наверху, служащий прикреплением спинных мышц. Однако даже позвоночник стал простой структурой. Нервная дуга, из которой она была выростом, обычно больше не сливается с позвоночным центром.
Шея короткая, и у производных видов уменьшения количества шейных позвонков. Короткая шея помещает близко к туловищу, обычно под небольшим наклоном к нему. Производные виды обычно также имеют уменьшенное количество спинных позвонков, общее количество пресакральных позвонков составляет от сорока до пятидесяти. Позвоночный столб мало дифференцирован. Приальной базтиоптеригии еще два крестцовых позвонка, но они не срослись. Формы раннего триаса имеют уплощенное в поперечном направлении основание хвоста с высокими шипами для волнообразного движения хвоста. Производные формы имеют более короткий хвост с характерным изгибом на конце; часть клиновидных позвонков, сама поддерживающая мясистую верхнюю часть хвостового плавника, вдавливающая конец хвоста в нижней части плавника.
производные виды больше не имеют поперечных отростков на своих позвонках - снова уникальное состояние для Амниота - парапофизарные и диапофизарные реберные суставы уменьшены до плоских позвонков, по крайней мере одна из граней установленных на телека. Количество граней может быть одним или двумя; их профиль может быть круглым или овальным. Их форма часто различается в зависимости от положения позвонка в колонне. Наличие двух граней на каждой стороне не означает, что само ребро двуглавое: даже в этом случае оно имеет единственную головку. Ребра обычно очень тонкие и имеют продольную канавку как на внутренней, так и на внешней стороне. Нижняя часть грудной клетки образована гастралией. Эти брюшные ребра имеют один центральный сегмент и один или два сегмента с каждой стороны. Они не сплавлены в настоящий пластрон. Обычно на дорсальном ребре присутствуют гастралии. Аппендикулярный скелет На этом образце, если смотреть снизу, то, что выглядит как грудина, на самом деле является сросшимися коракоидами На этом образце, если смотреть снизу, то, что выглядит как грудина, на самом деле является сросшимися коракоидами плечевым поясом ихтиозавров не сильно изменилось по сравнению с исходным состоянием. Некоторые базальные формы имеют форму топора или полумесяца лопатку или лопатку ; производные формы имеют удлиненный клинок, расположенный на более широком основании. Лопатка не срослась с коракоидом в скапулокоракоид, что указывает на то, что силы, действующие на плечевой пояс, были умеренными. Плечевой сустав расположен на границе между лопаткой и клювовидным отростком. Оба коракоида сливаются на общей средней линии. Форма коракоидов очень разнообразна, но обычно она довольно низкая. Верхняя часть плечевого пояса образована двумя тонкими ключицами, увенчанными центральными большими треугольными формами с базовой формой, маленькой и Т-образной у юрских видов. Грудные кости или грудины отсутствуют.  Ихтиозавр 'весло' (Береговый центр наследия Чармута ) Ихтиозавр 'весло' (Береговый центр наследия Чармута )Базальные формы имеют переднюю конечность, в некоторых деталях напоминают руки их наземных предков; локтевая кость и радиус удлинены и несколько разделены; запястья закруглены, что позволяет запястью вращаться; количество фаланг находится в пределах, показываемого наземными животными, собственно ихтиозавры, напротив, имеют переднюю конечность, которая полностью приспособлена к своей функции. Триасовые виды обычно имеют очень производный humerus, преобразованный в диск. Юрские виды, как правило, имеют более удлиненную форму плечевой кости с округлой головкой, узким стержнем и расширенным нижним концом. Локтевая кости всегда сильно уплощены, но могут иметь округлую форму. быть новыми для мед. У юрских форм больше нет пространства, межкостное пространство, между лучевой и локтевой костями. Часто последние кости постепенно переходят в нижние, дискообразные элементы - до четырех запястных костей, которые опять же мало отличаются по форме от пястных костей.  В этой руке Ophthalmosaurus icenius, появился дополнительный верхний ряд элементов, заканчивающийся наверху дополнительная кость нижней части руки. В этой руке Ophthalmosaurus icenius, появился дополнительный верхний ряд элементов, заканчивающийся наверху дополнительная кость нижней части руки. В строго производном состоянии показаны фаланги, небольшие дискообразные элементы, расположенные в последовательных рядах. Иногда количество пальцев уменьшается до двух. Это довольно распространенное явление в Tetrapoda. Однако уникальным для производных четвероногих является тот факт, что у некоторых видов наблюдается непатологическая полидактилия, когда количество пальцев превышает пять. У некоторых видов по 10 пальцев на руке. Эти пальцы, опять же, могут иметь увеличенное количество фаланг, до 30, явление, известное также из плезиозавров мозазавров и китообразных. Большое количество элементов позволяет флипперу иметь форму судна на подводных крыльях. Когда присутствует большое количество пальцев, их трудно определить. Обычно, что пальцы были добавлены как спереди, так и сзади, возможно, к сердцевине четырех исходных пальцев. Если пальцы складываются, часто увеличивается количество пястных и запястных костей; иногда присутствует даже лишний элемент нижнего рычага. Раньше ихтиозавры обычно подразделялись на «длинноперистые» и «латиперинговые» формы в зависимости от длинной или широкой передних ласт, но недавние исследования показали, что это не естественные группы; Клады ихтиозавров часто содержат виды с удлиненными передними конечностями и без них.
Таз ихтиозавра обычно довольно уменьшен. Три тазовые кости: подвздошная кость, седалищная кость и лобковая кость не срослись и часто не касаются друг друга. Кроме того, левая и правая тазовые стороны больше не соприкасаются; только у базальных форм еще есть крестцовые ребра, соединяющие подвздошную кость с позвоночником. Тазобедренный сустав изнутри не закрыт. Лобковая кость обычно не соединяется с седалищной костью позади нее; пространство между ними определено некоторыми исследователями как fenestra thyreoidea; другие исследователи отрицают применимость этого термина, данная общая рыхлую структуру таза. У некоторых более поздних видов лобковая кость и седалищная кость связаны, но в этом случае головка бедренной кости больше не сочленяется с тазобедренным суставом. Виды триаса имеют пластинчатые лобковые кости и седалищные кости; у более поздних видов эти элементы становятся удлиненными с узким стержнем и могут образовывать единый стержень.
Как правило, задние конечности короче передних и содержат меньшее количество элементов. Часто задний плавник составляет только половину длины переднего плавника. Бедренная кость короткая и широкая, часто с узкой талией и расширенным нижним концом. Большеберцовая, малоберцовая и плюсневые кости сливаются в мозаику из костных дисков, поддерживающих подводное крыло. Присутствуют от трех до шести пальцев ног. Фаланги пальцев ног также демонстрируют гиперфалангию; В исключительных случаях офтальмозавр показывает уменьшенное количество фаланг. Мягкие ткани Ихтиозавр Хольцмадена, у которого приготовитель обнаружил органические остатки на месте спинного плавника, но не смог их найти. для ласт. Ихтиозавр Хольцмадена, у которого приготовитель обнаружил органические остатки на месте спинного плавника, но не смог их найти. для ласт. На всех ранних реконструкциях ихтиозавров отсутствовали спинные плавники и хвостовые (хвостовые) двуустки, которые не поддерживались какой-либо твердой скелетной структурой, поэтому не сохранились во многих окаменелостях. Позвоночный столб поддерживает только нижнюю долю хвоста. В начале 1880-х годов были обнаружены первые очертания тела ихтиозавров. В 1881 году Ричард Оуэн сообщил об очертаниях тела ихтиозавра, показывающих хвостовые плавники из нижнеюрских пород в Барроу-апон-Соре, Англия. Другие хорошо сохранившиеся экземпляры с тех пор показали, что у некоторых более примитивных ихтиозавров, таких как образец Chaohusaurus geishanensis, хвостовая двуустка была развита слабо и имела только спинную долю хвоста, что делало хвост более похожим на весло. С годами видимость мочки хвоста у этого экземпляра исчезла.
Наличие спинных плавников у ихтиозавров было спорным. Прекрасно сохранившиеся образцы из Holzmaden Lagerstätten в Германии, найденные в конце 19 века, выявили дополнительные следы, обычно сохраненные черным цветом, контура всего тела, включая первые свидетельства дорсального плавники у ихтиозавров. Уникальные условия позволили сохранить эти очертания, которые, вероятно, состоят из бактериальных матов, а не остатков самих исходных тканей. В 1987 году утверждал, что, учитывая косвенный метод сохранения бактериями, эти очертания вряд ли надежно сохранились в каких-либо мелких деталях. Он пришел к выводу, что никаких настоящих спинных плавников обнаружено не было. После того, как смещенные кожные лоскуты тела изначально были ошибочно приняты за плавники, специалисты по обработке ископаемых позже стали ожидать наличия таких плавников и идентифицировали любое изменение цвета в соответствующем месте как спинной плавник или даже фальсифицировали такие структуры. Отсутствие спинного плавника также объясняет, почему ихтиозавры, в отличие от морских свиней, сохранили задние плавники, поскольку они были необходимы для стабильности. Другие исследователи отметили, что, хотя контуры могли быть заострены и сглажены специалистами по подготовке, потому что маты ископаемых бактерий обычно имеют нечеткие края, многие сохранившиеся спинные плавники, вероятно, были подлинными и, по крайней мере, в некоторой степени близки к истинному контуру тела. По крайней мере, один образец, R158 (в коллекции Paleontologiska Museet, Uppsala University ), показывает ожидаемые выцветшие края бактериального мата, поэтому он не был изменен подготовителями, но все еще сохраняет в целом тунца очертания тела, включая спинной плавник. В 1993 году Мартилль признал, что по крайней мере некоторые образцы спинных плавников являются подлинными.
Ископаемые образцы, сохранившие спинные плавники, также показали, что ласты были заостренными и часто намного шире, чем можно было бы предположить. Плавники поддерживались фиброзной тканью. В некоторых образцах видны четыре слоя коллагена, причем волокна покрывающих слоев пересекают слои нижнего коллагена.
В 2017 году на немецком сланце Posidonia Shale сообщалось об открытии 182,7 - позвонки Stenopterygius возрастом в миллион лет в карбонатном узле, все еще содержащие волокна коллагена, холестерин, тромбоциты, а также красные и белые кровяные тельца. Структуры не были бы окаменевшими, но представляли собой оригинальные органические ткани, биомолекулы которых можно было идентифицировать. Исключительная сохранность объяснялась защитной средой, создаваемой клубеньком. Обнаруженные эритроциты были от четверти до одной пятой по размеру, чем у современных млекопитающих. Это было бы приспособлением для улучшенного поглощения кислорода, в том числе ввиду низкого уровня кислорода во время тоара. Холестерин содержит изотоп с высоким содержанием углерода-13, что может указывать на более высокое положение в пищевой цепочке и в рационе из рыбы и головоногих.
В 2018 г. свидетельства жира был обнаружен с Stenopterygius. Кожа и окраска Ихтиозавр копролит Ихтиозавр копролит Как правило, сохранившиеся окаменелости предполагают, что кожа ихтиозавров была гладкой и эластичной, без чешуек. Однако эти останки - не отпечатки сами по себе, а очертания, сформированные в результате роста бактерий. В одном случае истинный отпечаток кожи был получен от образца Aegirosaurus, найденного в Solnhofen Plattenkalk, скалах, которые были способны сохранить даже мельчайшие детали. На этом образце, казалось, были видны крошечные чешуйки.
Окраску ихтиозавров трудно определить. В 1956 году Мэри Уайтар сообщила об обнаружении меланоцитов, пигментных клеток, в которых все еще должны были присутствовать красновато-коричневые пигментные гранулы, в образце кожи британской окаменелости, R 509. Традиционно считается, что ихтиозавры использовали затенение (темный сверху, светлый снизу), как акулы, пингвины и другие современные животные, служившие камуфляжем во время охоты. Этому противоречило открытие меланосом, структур, несущих черный меланин, в коже образца ихтиозавра YORYM 1993.338 Йоханом Линдгреном из Лундского университета. Был сделан вывод, что ихтиозавры, вероятно, имели однородную темную окраску для терморегуляции и для маскировки в глубокой воде во время охоты. Это контрастирует с мозазавридами и доисторическими кожистыми черепахами, которые, как было обнаружено, были затенены. Однако исследование 2015 года поставило под сомнение интерпретацию Линдгрен. В этом исследовании было отмечено, что базальный слой меланосом в коже повсеместно встречается в окраске рептилий, но не обязательно соответствует темноте. Другие структуры хроматофоров (такие как иридиофоры, ксантофоры и эритрофоры) влияют на окраску современных рептилий, но редко сохраняются или идентифицируются в окаменелостях. Таким образом, из-за неизвестного присутствия этих хроматофоров YORYM 1993.338 мог быть закрашенным, зеленым или иметь другие цвета или узоры. ГастролитыГастролиты, камни в желудке, которые могли способствовать пищеварению. или регулируемая плавучесть, лишь в нескольких случаях были обнаружены связанные со скелетами ихтиозавров, один раз с образцом Nannopterygius и второй раз с ископаемым Panjiangsaurus. Ихтиозавр копролиты, окаменелые фекалии, однако, очень распространены, их уже продала Мэри Эннинг. Палеобиология ЭкологияПомимо очевидного сходства с рыбами, ихтиозавры также имели общие черты развития с дельфинами, акулами-ламнидами и тунцом. Это придавало им в целом похожий вид, возможно, предполагало аналогичные уровни активности (включая терморегуляцию) и, по-видимому, помещало их в похожую экологическую нишу. Ихтиозавры не были преимущественно прибрежными животными; они также населяли открытый океан, что делало невозможным определение определенной области как места их происхождения. Кормление Temnodontosaurus acutirostris аммоноидей Temnodontosaurus acutirostris аммоноидей Ихтиозавры были плотоядными; они были настолько разнообразны по размеру и выживали так долго, что, вероятно, имели широкий спектр добычи. Виды с остроконечной мордой были приспособлены для хватания более мелких животных. МакГоуэн предположил, что формы с выступающей верхней челюстью в Eurhinosauria использовали свои заостренные морды, чтобы разрезать добычу, как это предполагалось для рыбы-меч.Наиболее часто сохраняемое содержимое кишечника ихтиозавров - это останки головоногих. Реже они питались рыбой и другими позвоночными, в том числе более мелкими ихтиозаврами. Большая форма триасового периода Thalattoarchon имел большие острые зубы и, вероятно, была способной убивать добычу своего размера, а также гималайзавр и несколько видов Temnodontosaurus. приспособления для убийства очень крупной добычи. Эти пищевые предпочтения подтверждены копролитами, действительно которые содержатки рыб и головоногих моллюсков. Еще одно подтверждение - окаменелое содержимое желудка. Бакленд в 1835 году описал присутствие в образце большой массы частично переваренных рыб, узнаваемых по их чешуе. Последующие исследования в 1968 году показали, что они принадлежали к роду рыб Pholidophorus, но также присутствовали клювы и присоски головоногих моллюсков. Такие твердые частицы пищи, по-видимому, задерживались в желудке и регулярно срыгивали. Также были съедены трупы утонувших животных: в 2003 г. Было сообщено, что у особи Platypterygius longmani, кроме рыб и черепахи, в желудке были кости наземной птицы.  Caypullisaurus подвергся нападению со стороны крокодиломорф дакозавр Caypullisaurus подвергся нападению со стороны крокодиломорф дакозавр Некоторые ранние ихтиозавры были дурофагами и имели плоские выпуклые зубы, приспособленные для дробления моллюсков. Таким образом, они ели бентос со дна мелководных морей. Другие виды, возможно, были присосками, засасывая животных в рот, быстро открывая их относительно короткие челюсти. Впервые это предполагалось для шонизавра, таким образом мог обеспечить постоянный запас пищи для своего огромного тела, а в 2011 году - для лианги Гуанлинзавр с короткой мордой. Однако в 2013 году исследование пришло к выводу, что подъязычная кость у ихтиозавров у основания языка была недостаточно окостеневшей, чтобы поддерживать движение всасывающего кормления, и предложили альтернативу, согласно которым такие виды были питающимися баранами, собирая пищу, постоянно плывя вперед с широко открытой пастью.
Типичные ихтиозавры имели очень большие глаза, защищенные костным кольцом, что предполагает, что они могли охотиться ночью или в большом количестве. глубина; единственные сохранившиеся животные такими же большими глазами - это гигантские и колоссальные кальмары. Таким образом, зрение, по-видимому, было одним из чувств, используемых во время охоты. Слух мог быть плохим, очень прочную форму стремени . Однако бороздки на нёбе предположить, что запах мог быть острым или даже могли присутствовать органы чувств.
Сами ихтиозавры служили пищей для других животных. В триасовый период их естественными хищниками в основном были акулы и другие ихтиозавры; в юрском периоде к ним присоединились крупные Плезиозаврия и морские Crocodylomorpha. Это снова подтверждается содержимым желудка: например, в 2009 г. был зарегистрирован образец плезиозавра с эмбрионом ихтиозавра в кишечнике. ПередвижениеУ ихтиозавров основная движущая сила обеспечивается боковым движением. тела. В ранней формех использовалось угловатое или угрейоподобное движение с волнообразными волнами всего туловища и хвоста. Обычно это считается неэффективным. Более поздние формы, такие как Parvipelvia, имели более короткие туловище и хвост, вероятно, использовали более эффективные или даже thunniform движения, в последнее третье тела, соответственно, хвостовая часть, только которых сгибается. Ствол у такого вида довольно жесткий.
Хвост был двудольным, нижняя доля поддерживалась хвостовым позвоночником, который был «изогнутым» вентрально, чтобы повторять контуры вентральной доли. У базальных видов хвостовой плавник довольно асимметричный или «гетероцеркальный». Асимметрия отличалась от таковой у акул тем, что нижняя доля была самой большой, а не верхней. Более производные формы имели почти вертикальный симметричный хвостовой плавник. Акулы используют свой асимметричный хвостовой плавник, чтобы компенсировать тот факт, что они обладают положительной плавучестью, тяжелее воды, за счет того, что давление, оказываемое хвостом вниз, заставляет тело в целом подниматься под углом. Таким образом, плавание вперед создаст необходимую подъемную силу. В 1973 году Макгоуэн пришел к выводу, что из-за ихтиозавров обратная асимметрия хвостового плавника по сравнению с акулами, они явно были плавучими, легче воды, что подтверждено отсутствием у них гастролитов и пахиостоза или плотной кости.. Хвост служил для удержания тела под опускающимся углом. Передние ласты будут главу для того, чтобы толкать переднюю часть тела дальше и контролировать . Однако в 1987 году Майкл А. Тейлор выдвинул альтернативную гипотезу: поскольку ихтиозавры могут изменить содержание своих легких, в отличие от акул (у отсутствует плавательный пузырь ), они также могут регулировать свою плавучесть. Таким образом, хвост в основном служил для нейтральной тяги, в то время как небольшие изменения плавучести стабилизировались небольшими изменениями угла наклона ласт. В 1992 году Макгоуэн принял эту точку зрения, указав, что акульи хвосты не являются хорошей аналогией хвостов ихтиозавров, которые имеют более узкие лопасти, более вертикальные и симметричные. Полученные хвостовые плавники ихтиозавра больше похожи на плавники тунца и указывают на сопоставимую способность выдерживать высокую крейсерскую скорость. Сравнительное исследование, проведенное Мотани в 2002 году, показало, что у современных животных маленькие доли хвостового плавника положительно коррелируют с высокой частотой биений. Современные исследователи в целом согласны с тем, что ихтиозавры обладали отрицательной плавучестью.  Несмотря на свои значительные размеры, плавники дельфина реки Амазонки в основном используются в качестве руля. Несмотря на свои значительные размеры, плавники дельфина реки Амазонки в основном используются в качестве руля. В 1994 году пришли к выводу, что ихтиозавры были самыми быстрыми морские рептилии. Их отношение длины к глубине составляло от трех до пяти, что является оптимальным числом для минимизации водонепроницаемости или сопротивления. Их гладкая кожа и обтекаемые тела предотвращали излишнюю турбулентность. Их степень преобразования энергии в поступательное движение приближается к величине дельфинов и составляет около 0,8. Ихтиозавры были бы на одну пятую быстрее плезиозавров, хотя половина разницы объяснялась предположением о 30% более высоком метаболизме ихтиозавров. Вместе, в рамках модели Массара, эти эффекты привели к крейсерской скорости чуть менее пяти километров в час. Однако в 2002 году Мотани исправил некоторые ошибки в формулах Массаре и изменил расчетную крейсерскую скорость до менее двух километров в час, что несколько ниже, чем у современных Cetacea. Однако, поскольку оценки скорости плезиозавров и мозазаврид также были пересмотрены в сторону уменьшения, ихтиозавры сохранили свое относительное положение.
Ихтиозавры имели плавниковые конечности разной относительной длины. Стандартная интерпретация состоит в том, что они, вместе со спинным и хвостовым плавниками, использовались в качестве управляющих поверхностей для курсовой устойчивости, управления рысканием и для стабилизации тангажа и крена, а не движения. Однако в 80-е годы немецкий палеонтолог предложил альтернативную модель. Изучив летающие движения передних конечностей плезиозавров, он предположил, что по крайней мере те ихтиозавры, у которых были длинные ласты, использовали их для мощного движущего движения, перемещая их вверх и вниз. Это объяснило бы не дегенерированный плечевой пояс и эволюцию костей рук, чей идеальный профиль на подводных крыльях был бы бесполезен, если бы он не использовался функционально. Он думал, что обнаружил современные аналоги у двоякодышащей рыбы Квинсленда и дельфина реки Амазонки, которые, как он предполагал, также использовали свои длинные плавники для движения. Рисс изложил эту гипотезу в серии статей. Эта альтернативная интерпретация обычно не принималась другими работниками. В 1998 году Даррен Нейш указал, что двоякодышащие и речной дельфин на самом деле не используют свои плавники таким образом, а, например, современный горбатый кит имеет очень длинные передние ласты, поддерживаемые мозаикой из костей, но, тем не менее, в основном они служат рулями. В 2013 году исследование пришло к выводу, что широкие ласты ихтиозавра, как и у Platyptergygius, использовались не для движения, а в качестве контрольной поверхности. Дайвинг Темнодонтозавр имел самые большие глаза среди всех известных позвоночных, что свидетельствует о хорошем способность нырять Темнодонтозавр имел самые большие глаза среди всех известных позвоночных, что свидетельствует о хорошем способность нырять Многие из ныне живущих морских позвоночных, дышащих легкими, способны к глубокому нырянию. Есть некоторые указания на способность ихтиозавров нырять. Быстрый подъем с большей глубины может вызвать декомпрессионную болезнь. Получившаяся кость некроз был хорошо документирован у ихтиозавров юрского и мелового периодов, где он присутствует в 15% и 18% экземпляров, соответственно, но редко встречается у видов триаса. Это могло быть признаком того, что базальные формы ныряли не так глубоко, но также могло быть объяснено более сильным давлением хищников в более поздние эпохи. Однако последней возможности противоречит тот факт, что у современных животных ущерб вызван не ограниченным числом инцидентов быстрого восхождения, а этим накоплением не обесценивающей дегенерации во время обычного ныряния.
Глазами ихтиоз предоставляют дополнительные сертификаты того, что среди позвоночных они абсолютно известны как самые крупные. Современные морские леопарды могут нырять на глубину до 1 км, охотясь на месте. Мотани предположил, что ихтиозавры с их большими глазами должны были иметь возможность получить еще большие глубин. Темнодонтозавр с глазами размером пять сантиметров, двадцать, все еще мог видеть на глубине 1600 метров. На такой глубине такие глаза были бы особенно полезны для просмотра крупных объектов. Более поздние виды, такие как Ophthalmosaurus, имели относительно большие глаза, что снова указывает на то, что способность нырять была лучше в позднеюрских и мых формах. МетаболизмПодобно современным китообразным, таким как киты и дельфины, ихтиозавры дышали воздухом. Киты и дельфины - млекопитающие и теплокровные. Об ихтиозаврах традиционно считалось хладнокровным, т. Е. рептилиями. Однако с 1970-х годов многие доминирующие группы рептилий мезозоя, такие как тероподы динозавры, птерозавры и плезиозавры, считались теплыми. - кровавые, так как это предлагает элегантное объяснение их доминирования. Имеются некоторые прямые доказательства того, что ихтиозавры тоже могли быть эндотермическими. В 1990 г. Вивиан де Баффрениль опубликовала гистологическое исследование, в котором указывалось, что ихтиозавры обладали фиброламеллярной структурой кости, как и у теплокровных животных в целом, характеризующейся быстрым ростом и сильным васкуляризация. Эти черты уже проявляются у видов раннего триаса. В 2012 году сообщалось, что даже самая базальная форма Utatsusaurus этот тип кости, что указывает на то, что предкитиозавров уже были теплокровными. Дополнительным прямым доказательством высокого метаболизма является соотношение изотопов кислорода в зубах, которое указывает на температуру тела от 35 до 39 ° C, что примерно на 20 ° выше, чем в окружающей морской воде.. Жир соответствует теплокровности, поскольку изолирующие свойства требуют, чтобы животное вырабатывало собственное тепло.
Косвенным доказательством эндотермии служит форма тела производных ихтиозавров, которые с коротким хвостом и вертикальным плавательным движением кажутсяальной скоростью, которая может быть поддержана полностью теплокровной скоростью: все ныне живущие животные, плавающими таким образом, либо, как акулы и тунцы, высокая температура в ядре своего тела. Этот аргумент не распространяется на базальные формы с более угрюмым телом и волнообразными плавательными движениями. В 1996 году их заставляет воздух из легких, предположил, что они подвержены уязвимому ограничению Карриера, ограничению рептильного дыхания, которое в 1987 году годузывалось: волнообразное движение из и, таким образом, не позволяет им дышать во время движения. Коуэн предположил, что ихтиозавры преодолели бы проблему с помощью морских свиней: постоянные выпрыгивания из воды позволили бы им глотать свежего воздуха во время прыжка. Кэрриера неприменимо из-за их жестких тел, кажется, подтверждено их хорошей способностью нырять, подразумевая эффективное дыхание и систему хранения кислорода, другие исследователи имели тенденцию предполагать, что, по крайней мере, для производных ихтиозавров ограничение. Для этих видов морских свиней не было необходимости. Тем не менее, ихтиозавры часто всплывали, чтобы дышать, вероятно, слегка наклонив голову, чтобы вдохнуть воздух, из-за более низкого положения ноздрей по сравнению с дельфинами. Размножение Chaohusaurus с тремя молодыми особями Chaohusaurus с тремя молодыми особями Ихтиозавры были живородящими, то есть рожали живых детенышей, а не откладывали яйца. Хотя были рептилиями и произошли от яйцекладущих яйцекладущих предков, живорождение не так неожиданно, как кажется на первый взгляд. Дышащие воздухом морские существа должны либо выйти на берег, чтобы отложить яйца, как черепахи и некоторые морские змеи, либо родить живых детенышей в поверхностных водах, например киты и дельфины. Учитывая их обтекаемые и поперечно сплюснутые тела, хорошо приспособленные для быстрого плавания, ихтиозаврам было бы трудно, если не невозможно, передвигаться по суше достаточно далеко, чтобы откладывать яйца. Это подтвердилось уже 9 декабря 1845 года, когда натуралист сообщил о небольшом эмбрионе в окаменелости Ichthyosaurus communis. Эмбрион длиной одиннадцать сантиметров помещен в родовые пути его матери длиной два с половиной метра, головой назад. На основании окаменелостей Пирс пришел к выводу, что ихтиозавры должны быть живородящими.  Деталь самки Stenopterygius с большим плодов в животе, один из которых был изгнан. Деталь самки Stenopterygius с большим плодов в животе, один из которых был изгнан. Позже из отложений Хольцмадена были обнаружены многочисленные окаменелости взрослых особей. найдено плодов. В 1880 году Гарри Говьер Сили, экспервший специальный британский пантологический комитет, из проблем воспроизводства ихтиозавров, пришел к выводу, что рождение происходящего в воде и что окаменелости, внесение зародыши в родовых путях, вероятно, случаев преждевременной смерти молодь, вызывая гибель материнского животного. Было проведено сравнение с дельфинами и китами, детенышей которых должны рождатьсяом вперед, чтобы не утонуть; если ребенок рождается головой вперед, он умирает вместе с матерью, если труп застревает в родовых путях. Однако альтернативное объяснение заключается в том, что такие окаменелости на самом деле, умершие по другим причинам во время беременности, после чего газы разложения вытеснили зародыши головой. В 2014 году в исследовании сообщалось об обнаружении окаменелой самки Chaohusaurus, который умерла при родах трех новорожденных. Двое уже были изгнаны, а третий находился в родовых путях. Окаменелость также свидетельствует о том, что ранние ихтиозавры также рождались головой вперед, возможно, в отличие от более поздних родов. Так как Chaohusaurus является ихтиозавром на очень низком уровне - ранее наиболее базовым родом, зародыши которого были известны, был Mixosaurus - это открытие предполагает, что самые ранние наземные предки ихтиозавров уже были живородящими . По сравнению с плацентарными млекопитающими или плезиозаврами, плоды ихтиозавров, как правило, очень малы, а их количество в помете часто велико. У одной самки Stenopterygius было идентифицировано семь, у другой - одиннадцать. Длина плода составляет не более четверти длины материнского животного. Молодые особи имеют примерно такие же пропорции тела, как и взрослые особи. Основные онтогенетические изменения во время роста состоят в слиянии и прочности скелетных элементов.
Крокодилы, большинство морских черепах и некоторые ящерицы контролируют пол потомства, манипулируют температурной окружающей средой для разных яиц; т.е. у них нет различного пола хромосом. Живородящие рептилии не регулируют посредством температуры инкубации. Исследование 2009 года, в котором были изучены 94 живых вида рептилий, птиц и млекопитающих, показало, что генетический контроль пола имеет значение решающее для живорождения. Был сделан вывод, что у морских рептилий такой контроль предшествовал живорождению и являлся адаптацией к стабильному морскому климату в прибрежных регионах. Генетика, вероятно, контролируется пол у ихтиозавров, мозазавров и плезиозавров. Социальное поведение и интеллектИхтиозавры часто жили стадами или охотничьими группами. Имеется мало свидетельств о природе ихтиозавров социального поведения. Существуют некоторые признаки наличия уровня полового диморфизма. Скелеты евринозавра и шастазавра имеют два морфотипа. Особи с более длинной мордой, большими глазами, более новыми туловищами, более коротким хвостом и более дополнительными фалангами могли представить самок; более длинный ствол мог обеспечить место для эмбрионов.
Обычно мозг имеет ограниченный размер и удлиненную форму, как у современных хладнокровных рептилий. Однако в 1973 году Макгоуэн, изученный естественный эндокаст хорошо сохранившегося образца, был указан, что конечный мозг не очень маленьким. Зрительные мочки были большими, как и следовало ожидать по размеру глаза. Обонятельные доли были, хотя и не сильно, но хорошо дифференцированными; то же самое верно и в отношении мозжечка. Патологииизвестно, хотя, что окаменелости, свидетельствующие о поведении ихтиозавра, остаются редкими, что одна окаменелость ихтиозавра постоянно укусила область морды. Обнаруженные в Австралии и проанализированные и др. В 2011 году измерения ран показывают, что следы укусов были нанесены другими ихтиозавром, вероятно, того же вида, что является вероятным случаем укуса лица во время конфликта. Раны показывают признаки заживления в виде роста костей, а это означает, что жертва пережила нападение. Другой, очень большой ихтиозавр длиной около девяти метров был найден на Свальбарде ; он был почти готов, за исключением хвоста. Изучение находки показало, что во время охоты на аммонитов (о чем свидетельствует доказина аммонита в области горла), ихтиозавр попал в засаду и подвергся нападению, вероятно, со стороны плиозаврида (известного из той же среды обитания), который разорвал его хвост. Затем ихтиозавр опустился на глубину, утонул и, в итоге, окаменел в глубокой воде. Находка была представлена в специальном разделе National Geographic «Смерть морского чудовища». Геологические образования Ниже представлен список геологических образований, в которых были найдены окаменелости ихтиозавра: | Имя | Возраст | Местоположение | Примечания |
|---|
Формация Агардфьеллет | поздний Титон | | Криоптеригиус, Янусавр, Палвенния | Формация Антимонио | конец Карнийский период | | Shastasaurus pacificus, Toretocnemus californicus | Формация Besano | Средний триас | | Безанозавр, Cymbospondylus buchseri, Mikadocephalus, Mixosaurus Cornalianus, Mixosaurus kuhnschnyderi, Phalarodon fraasi, Phalarodon major, Tholodus, Wimanius | Блю Лиас | Синемуриан | | Ихтиозавр communis, Leptonectes tenuirostris, Temnodontosaurus Platyodon | Clearwater Formati на | ранний альбский | | атабасказавр | формация Фавре () | анисианский | | Cymbospondylus nichollsi, Phalarodon callawayi, Phalarodon fraasi, Thalattoarchon | францисканская формация | | | | формация Guanling | Anisian | | Barracudasauroides, Contectopalatus, Xinminosaurus | Hosselkus Limestone | поздний Карнийский | | Калифорнозавр, Shastasaurus pacificus, Toretocnemus californicus, Toretocnemus zitteli | | Оленекян | | Chaohusaurus zhangjiawanensis | | Форма Катрол | Киммеридж | | Индетерминантные «ихтиозавры» | | Ранний триас | | Тайзавр | Киммеридж Клей | Киммеридж | | Brachypterygius extremus, Нанноптеригиус | Формация реки Лун | ранний Альбский | | Майаспондил | Форма Лос-Моллес | ранняя байосская | | Чакаикозавр, Моллес aurus | Нижний Гринсанд | | | | Формация Лунинг | поздний Карнийский | | Shonisaurus popularis | Muschelkalk | Средний триас | | Contectopalatus, Cymbospondylus germanicus, Cymbospondylus parvus, Omphalosaurus peyeri, Omphalosaurus wolfi, Phalarodon major, Фантомозавр, Толодус | | поздний Оленекский | | Chaohusaurus geishanensis | Ньялам | Норианский | | Гималайазавр | | ранний Аален | | Stenopterygius aaleniensis | (верхний) | Оленекский | | Utatsusaurus | Oxford Clay | Келловейский | | Ophthalmosaurus icenicus | Формация Паха | Аптиан | | Muiscasaurus, Platypterygius sachicarum | Формация Пардонет | средний Норианский | | Каллавайя, Hudsonelpidia, Macgowania, Shonisaurus / Shastasaurus sikkannie nsis | Posidonia Shale | ранний тоарский | | Eurhinosaurus longirostris, Hauffiopteryx, Stenopterygius quadriscissus, Stenopterygius трисцисс, стеноптеригиус унитер, суеволевиафан 58>дезинтегер, суеволевиафан целое число | формация Прида () | анисианский | | Cymbospondylus petrinus, Omphalosaurus nettarhynchus, Omphalosaurus nevadanus, Phalarodon fraasi | Формация Рингнес | Оксфорд - Киммеридж | | Arthropterygius | Солнхофенский известняк | Титонский | | Эгирозавр | Спитон Клей | Готерив | | Acamptonectes | | поздний Оленек 881>Норвегия | Гриппия, Исфьордозавр, Omphalosaurus merriami, Pessopteryx, Quasianosteosaurus | Strawberry Bank, Ilminster | ранний тоарский | | Hauffiopteryx, Stenopterygius трисцисс | горная формация серы | поздний оленекский - анизийский | | гулозавр, Парв инататор, Phalarodon fraasi, Utatsusaurus | Sundance Formation | | | | Tschermakfjellet Formation | ладинский - карнийский | | Cymbospondylus sp., Mikadocephalus, Phalarodon callawayi, Phalarodon fraasi | Vaca Muerta | Tithonian | | Caypullisaurus | Формация Сяова | Карнийский | | Гуаньлинзавр, Гуйчжоуичтиозавр tangae, Гуйчжоуичтиозавр Волонггангенс, Цианихтиозавр известный как член Wayao. |
См. Также Ссылки Источники Внешние ссылки  | На Wikimedia Commons есть материалы, связанные с Ихтиозаврией. |
Последняя правка сделана 2021-05-23 10:17:52
Содержание доступно по лицензии CC BY-SA 3.0 (если не указано иное).
Обратная связь: support@alphapedia.ru

|


 Ichthyosauria Плезиозавр, автор Эдуард Рио, 1863. (Это старое изображение плезиозавра, поднимающего голову, неточно и физически невозможно )
Ichthyosauria Плезиозавр, автор Эдуард Рио, 1863. (Это старое изображение плезиозавра, поднимающего голову, неточно и физически невозможно ) Череп, найденный Джозефом Эннингом в 1811 году
Череп, найденный Джозефом Эннингом в 1811 году 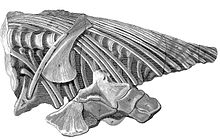 Туловище, найденное Мэри Эннинг в 1812 году
Туловище, найденное Мэри Эннинг в 1812 году  «Профессор Ихтиозавр» показывает своим ученикам череп вымерший человек, карикатура Генри Де ла Беша
«Профессор Ихтиозавр» показывает своим ученикам череп вымерший человек, карикатура Генри Де ла Беша  Образцы Хокинса до сих пор являются экспонатами Музея естественной истории
Образцы Хокинса до сих пор являются экспонатами Музея естественной истории  Типичные Holzmaden окаменелости: взрослые и молодые Stenopterygius quadriscissus
Типичные Holzmaden окаменелости: взрослые и молодые Stenopterygius quadriscissus  Драгоценный опал, заменяющий хребет ихтиозавра, экспонат, Южно-Австралийский музей
Драгоценный опал, заменяющий хребет ихтиозавра, экспонат, Южно-Австралийский музей  Grippia longirostris из раннего триаса на Шпицбергене уже было хорошо приспособлены к водному образу жизни.
Grippia longirostris из раннего триаса на Шпицбергене уже было хорошо приспособлены к водному образу жизни.  Hupehsuchus
Hupehsuchus 

 Стеноптеригиус напоминал современного дельфина.
Стеноптеригиус напоминал современного дельфина.  Многие линии ихтиозавров продолжились и в меловом периоде.
Многие линии ихтиозавров продолжились и в меловом периоде.  Реставрация
Реставрация  CGI-реставрация
CGI-реставрация  Примеры отличительных черт, присущих как дельфинам, так и производным ихтиоптеригам.
Примеры отличительных черт, присущих как дельфинам, так и производным ихтиоптеригам.  Череп
Череп  Позвонок ихтиозавра из
Позвонок ихтиозавра из  На этом образце, если смотреть снизу, то, что выглядит как грудина, на самом деле является сросшимися
На этом образце, если смотреть снизу, то, что выглядит как грудина, на самом деле является сросшимися  Ихтиозавр 'весло' (
Ихтиозавр 'весло' ( В этой руке
В этой руке  Ихтиозавр Хольцмадена, у которого приготовитель обнаружил органические остатки на месте спинного плавника, но не смог их найти. для ласт.
Ихтиозавр Хольцмадена, у которого приготовитель обнаружил органические остатки на месте спинного плавника, но не смог их найти. для ласт.  Ихтиозавр
Ихтиозавр 

 Несмотря на свои значительные размеры, плавники
Несмотря на свои значительные размеры, плавники  Темнодонтозавр имел самые большие глаза среди всех известных позвоночных, что свидетельствует о хорошем способность нырять
Темнодонтозавр имел самые большие глаза среди всех известных позвоночных, что свидетельствует о хорошем способность нырять  Chaohusaurus с тремя молодыми особями
Chaohusaurus с тремя молодыми особями  Деталь самки Stenopterygius с большим плодов в животе, один из которых был изгнан.
Деталь самки Stenopterygius с большим плодов в животе, один из которых был изгнан.