
Войти
| Cephalopod. Временной диапазон: Поздний кембрий - Настоящее время До Ꞓ O S D C P T J K Pg N | |
|---|---|
 | |
| рифовый кальмар. (Sepioteuthis lessoniana) | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Mollusca |
| Класс: | Cephalopoda. Cuvier, 1797 |
| Подклассы | |
A cephalopod () - любой представитель моллюсков класса Cephalopoda (греч. множественное число κεφαλόποδες, kephalópodes; -feet "), например кальмар, осьминог, каракатица или наутилус. Эти исключительно морские животные характеризуются двусторонней симметрией тела, выдающейся головой и набором рук или щупалец (мышечные гидростаты ). модифицированный из примитивной ноги моллюска. Рыбаки иногда называют головоногих моллюсков «чернилами », имея в виду их обычную способность брызгать чернилами. Изучение головоногих моллюсков - это раздел малакологии, известный как тевтология.
. Головоногие моллюски стали доминирующими в ордовикский период, представленные примитивными наутилоидами. Класс теперь содержит два, только отдаленно связанных, существующих подкласса : Coleoidea, в который входят осьминоги, кальмары и каракатицы. ; и Nautiloidea, представленный Nautilus и Allonautilus. У Coleoidea раковина моллюска интернализована или отсутствует, тогда как у Nautiloidea внешняя раковина остается. Идентифицировано около 800 живых видов головоногих моллюсков. Два важных вымерших таксона - это Ammonoidea (аммониты) и Belemnoidea (белемниты).

 Слева: Пара Sepia officinalis на мелководье. Справа: Benthoctopus sp. на подводной горе Дэвидсон на глубине 2422 м
Слева: Пара Sepia officinalis на мелководье. Справа: Benthoctopus sp. на подводной горе Дэвидсон на глубине 2422 м Существует более 800 существующих видов головоногих моллюсков, хотя новые виды продолжают описываться. Описано около 11000 вымерших таксонов, хотя мягкая природа головоногих моллюсков означает, что их нелегко окаменелить.
Головоногие моллюски встречаются во всех океанах Земли. Ни один из них не переносит пресноводный, но короткий кальмар, Lolliguncula brevis, обнаруженный в Чесапикском заливе, является заметным частичным исключением, поскольку он терпит солоноватая вода. Считается, что головоногие не могут жить в пресной воде из-за множества биохимических ограничений, и за время своего существования более 400 миллионов лет никогда не осмеливались проникать в полностью пресноводные среды обитания.
Головоногие моллюски занимают большую часть глубины океана, начиная с моря. бездонная равнина до поверхности моря. Их разнообразие наиболее велико вблизи экватора (~ 40 видов, пойманных в сети на 11 ° с.ш. по результатам исследования разнообразия) и уменьшается к полюсам (~ 5 видов, пойманных на 60 ° с.ш.).

 Слева: осьминог, открывающий контейнер с завинчивающейся крышкой. Справа: кальмар гавайский бобтейл, сколопы Euprymna, зарываясь в песок, оставляющий открытыми только глаза
Слева: осьминог, открывающий контейнер с завинчивающейся крышкой. Справа: кальмар гавайский бобтейл, сколопы Euprymna, зарываясь в песок, оставляющий открытыми только глаза Головоногие моллюски считаются наиболее умными из беспозвоночных, у них хорошо развиты чувства и большой мозг (больше, чем у >брюхоногие моллюски ). нервная система головоногих моллюсков является наиболее сложной из беспозвоночных, и соотношение их массы мозга и массы тела находится между эндотермическими и экзотермическими позвоночными. Известно также, что головоногие моллюски в неволе вылезают из своих аквариумов, маневрируют на расстоянии от пола лаборатории, заходят в другой аквариум, чтобы поесть крабов, и возвращаются в свой собственный аквариум.
Мозг защищен в хрящевой череп. Гигантские нервные волокна головоногих мантии в течение многих лет широко использовались в качестве экспериментального материала в нейрофизиологии ; их большой диаметр (из-за отсутствия миелинизации ) делает их относительно простыми для изучения по сравнению с другими животными.
Многие головоногие моллюски - существа социальные; когда они изолированы от себе подобных, некоторые виды наблюдались стадами с рыбой.
Некоторые головоногие моллюски способны летать по воздуху на расстояние до 50 м. Хотя головоногие моллюски не особенно аэродинамичны, они достигают этих впечатляющих расстояний с помощью реактивного движения; вода продолжает выходить из воронки, пока организм находится в воздухе. Животные расправляют плавники и щупальца, образуя крылья, и активно контролируют подъемную силу с помощью положения тела. У одного вида, Todarodes pacificus, было замечено распространение щупалец в форме плоского веера со слизистой пленкой между отдельными щупальцами, в то время как у другого, Sepioteuthis sepioidea, было обнаружено, что щупальца складываются в круговое расположение.
Головоногие обладают развитым зрением, могут обнаруживать гравитацию с помощью статоцист и имеют множество химических органов чувств. Осьминоги используют свои руки для исследования окружающей среды и могут использовать их для восприятия глубины.
 Примитивный глаз наутилус работает так же, как камера-обскура.
Примитивный глаз наутилус работает так же, как камера-обскура. Воспроизвести медиа W-образный зрачок каракатицы расширяется при выключении света.
Воспроизвести медиа W-образный зрачок каракатицы расширяется при выключении света. Большинство головоногих моллюсков полагаются на зрение, чтобы обнаруживать хищников и добычу и общаться друг с другом. Следовательно, зрение головоногих моллюсков остро: обучающие эксперименты показали, что обыкновенный осьминог может различать яркость, размер, форму и горизонтальную или вертикальную ориентацию объектов. Морфологическая конструкция дает глазам головоногих моллюсков такие же характеристики, как и у акул; однако их конструкция отличается, поскольку у головоногих моллюсков отсутствует роговица, а сетчатка вывернута. Глаза головоногих также чувствительны к плоскости поляризации света. В отличие от многих других головоногих моллюсков, наутилусы не обладают хорошим зрением; их глазная структура сильно развита, но у них отсутствует прочная линза. У них есть простое "отверстие ", через которое может проходить вода. Считается, что вместо зрения животное использует обоняние в качестве основного чувства для поиска пищи, а также для поиска или идентификации потенциальных партнеров.
 Каракатица с W-образными зрачками, которые могут помочь им различать цвета.
Каракатица с W-образными зрачками, которые могут помочь им различать цвета. Удивительно, учитывая их способность изменять цвет, все осьминоги и большинство головоногих считаются дальтониками. Колеидные головоногие моллюски (осьминоги, кальмары, каракатицы) имеют один тип фоторецепторов и не имеют возможности определять цвет, сравнивая интенсивность обнаруженных фотонов по нескольким спектральным каналам. Когда маскируют себя, они используют свои хроматофоры для изменения яркости и рисунка в соответствии с видимым фоном, но их способность соответствовать определенному цвету фона может исходить от таких ячеек, как иридофоры и лейкофоры, отражающие свет из окружающей среды. Они также производят визуальные пигменты по всему телу и могут ощущать уровни света непосредственно от своего тела. Свидетельства цветового зрения были обнаружены у искрящегося кальмара энопа (Watasenia scintillans), у которого цветное зрение достигается за счет использования трех различных молекул сетчатки : A1, наиболее чувствителен к зелено-синему (484 нм); A2 - сине-зеленым (500 нм) и A4 - синим (470 нм), которые связываются с его опсином.
. В 2015 году был описан новый механизм спектральной дискриминации у головоногих моллюсков. Это основано на использовании хроматической аберрации (зависимость фокусного расстояния от длины волны). Численное моделирование показывает, что хроматическая аберрация может дать полезную хроматическую информацию через зависимость резкости изображения от аккомодации. Необычная внеосевая щель и кольцевая форма зрачка у головоногих моллюсков усиливают эту способность.
В 2015 году были опубликованы молекулярные данные, указывающие на то, что хроматофоры головоногих моллюсков светочувствительны; Полимеразная цепная реакция с обратной транскрипцией (ОТ-ПЦР) выявила транскрипты, кодирующие родопсин, и в сетчатке и коже прибрежного длинноперого кальмара (Doryteuthis pealeii) и каракатица обыкновенная (Sepia officinalis) и каракатица широкая (Sepia latimanus ). Авторы утверждают, что это первое доказательство того, что кожные ткани головоногих моллюсков могут обладать необходимой комбинацией молекул для ответа на свет.
Было показано, что некоторые кальмары улавливают звук, используя свои statocysts.
 Эта ширококлювая каракатица (Sepia latimanus) может меняться с камуфляжно-коричневого и коричневого (вверху) на желтый с темными бликами (внизу) менее чем за секунду.
Эта ширококлювая каракатица (Sepia latimanus) может меняться с камуфляжно-коричневого и коричневого (вверху) на желтый с темными бликами (внизу) менее чем за секунду. Большинство головоногих моллюсков обладают набором компонентов кожи, взаимодействующих со светом. Они могут включать иридофоры, лейкофоры, хроматофоры и (у некоторых видов) фотофоры. Хроматофоры - это цветные пигментные клетки, которые расширяются и сжимаются в соответствии с цветом и узором, которые они могут использовать в потрясающем множестве мод. Помимо маскировки своим фоном, некоторые головоногие моллюски биолюминесцируют, светят вниз, чтобы скрыть свои тени от любых хищников, которые могут скрываться внизу. биолюминесценция производится бактериальными симбионтами; головоногие моллюски-хозяева способны обнаруживать свет, излучаемый этими организмами. Биолюминесценция также может использоваться для соблазнения добычи, а некоторые виды используют красочные дисплеи, чтобы произвести впечатление на товарищей, напугать хищников или даже общаться друг с другом.
Головоногие моллюски могут менять свой цвет и узор за миллисекунды, будь то для сигнализации (как внутри вида, так и для предупреждения ) или активный камуфляж, поскольку их хроматофоры расширены или сжаты. Хотя изменения цвета, по-видимому, в основном зависят от информации, получаемой от зрения, есть свидетельства того, что клетки кожи, в частности хроматофоры, могут обнаруживать свет и приспосабливаться к условиям освещения независимо от глаз. Головоногие могут использовать хроматофоры как мышцы, поэтому они могут менять цвет своей кожи так же быстро, как и они. Окраска обычно сильнее у прибрежных видов, чем у видов, живущих в открытом океане, чьи функции обычно ограничиваются разрушительной маскировкой. Большинство осьминогов имитируют избранные структуры в своем поле зрения, а не становятся составным цветом своего полного фона.
Доказательства первоначальной окраски были обнаружены в окаменелостях головоногих моллюсков, датируемых еще силурийским периодом ; у этих ортоконических особей были концентрические полосы, которые, как считается, служили камуфляжем. Девонские головоногие моллюски имеют более сложные цветовые узоры, функция которых неизвестна.
За исключением Nautilidae и видов осьминогов, принадлежащих к подотряд Циррина, все известные головоногие моллюски имеют чернильный мешок, который можно использовать для изгнания облака темных чернил, чтобы сбить с толку хищников. Этот мешок представляет собой мышечную сумку, которая возникла как продолжение задней кишки. Он находится под кишечником и открывается в задний проход, в который можно впрыснуть его содержимое - почти чистый меланин ; его близость к основанию воронки означает, что чернила могут распространяться за счет выбрасываемой воды, поскольку головоногие моллюски используют свою реактивную тягу. Выброшенное облако меланина при изгнании обычно смешивается с слизью, образующейся в другом месте мантии, и поэтому образует густое облако, что приводит к ухудшению зрения (и, возможно, химиосенсорных функций) хищника, как у дымовая завеса. Однако наблюдалось более изощренное поведение, при котором головоногие выпускают облако с большим содержанием слизи, которое приблизительно напоминает головоногих, выпустивших его (эта приманка упоминается как Псевдоморф ). Эта стратегия часто приводит к нападению хищника на псевдоморфа, а не на его быстро убегающую добычу. Для получения дополнительной информации см. Чернильное поведение.
Чернильный мешок головоногих моллюсков дал общее название «чернильная рыба», ранее называемая чернильной рыбой.
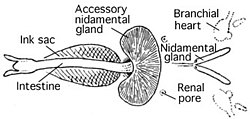 Внутренности Chtenopteryx sicula
Внутренности Chtenopteryx sicula  Внутренности Ocythoe tuberculata
Внутренности Ocythoe tuberculata Головоногие моллюски - единственные моллюски с закрытой системой кровообращения. Колеоиды имеют две жабры сердца (также известные как жаберные сердца ), которые перемещают кровь через капилляры жабр. Затем одно системное сердце перекачивает насыщенную кислородом кровь через остальное тело.
Как и большинство моллюсков, головоногие моллюски используют гемоцианин, содержащий медь белок, а не гемоглобин <352.>, чтобы транспортировать кислород. В результате их кровь становится бесцветной при деоксигенации и становится синей при контакте с воздухом.
Головоногие моллюски обмениваются газами с морской водой, проталкивая воду через жабры, прикрепленные к крыше. организма. Вода поступает в мантийную полость снаружи жабр, и вход в мантийную полость закрывается. Когда мантия сжимается, вода выталкивается через жабры, которые находятся между полостью мантии и воронкой. Вытеснение воды через воронку может быть использовано для приведения в действие водометного движения. Жабры, которые намного эффективнее, чем у других моллюсков, прикрепляются к вентральной поверхности мантийной полости. Размер жабр зависит от образа жизни. Чтобы достичь высоких скоростей, жабры должны быть небольшого размера - вода будет проходить через них быстро, когда потребуется энергия, компенсируя их небольшой размер. Однако организмы, которые проводят большую часть своего времени, медленно перемещаясь по дну, естественным образом не пропускают через свою полость много воды для передвижения; таким образом, у них есть более крупные жабры, а также сложные системы, обеспечивающие постоянную промывку воды через жабры, даже когда организм неподвижен. Водный поток контролируется сокращениями радиальных и круговых мышц мантийной полости.
Жабры головоногих моллюсков поддерживаются скелетом из прочных волокнистых белков; Отсутствие мукополисахаридов отличает этот матрикс от хряща. Считается, что жабры также участвуют в выделении, при этом NH 4 заменяется K из морской воды.
 Осьминоги плывут головой вперед, с вытянутыми сзади руками
Осьминоги плывут головой вперед, с вытянутыми сзади руками Хотя большинство головоногих моллюсков могут двигаться с помощью реактивной тяги, это очень энергоемкий способ передвижения по сравнению с движением хвоста, используемым рыбами. Эффективность пропеллера водомет (то есть КПД Фруда ) выше, чем ракеты. Относительная эффективность реактивного движения дополнительно снижается по мере увеличения размера животного; paralarvae намного более эффективны, чем молодые и взрослые особи. Начиная с палеозойской эры, поскольку конкуренция с рыбой создавала среду, в которой эффективное движение было решающим для выживания, реактивное движение отошло на второй план с плавниками и щупальца используются для поддержания постоянной скорости. Хотя реактивное движение никогда не является единственным способом передвижения, движение с остановкой и запуском, обеспечиваемое реактивными двигателями, по-прежнему полезно для обеспечения всплесков высокой скорости - не в последнюю очередь при поимке добычи или избегании хищников. Действительно, это делает головоногих моллюсков самыми быстрыми морскими беспозвоночными, и они могут опережать большинство рыб. Струя дополнена движением плавников; у кальмара ласты хлопают каждый раз при выпуске струи, усиливая тягу; затем они вытягиваются между струями (предположительно, чтобы избежать затопления). Кислородная вода попадает в полость мантии к жабрам, и в результате мышечного сокращения этой полости отработанная вода выталкивается через гипоном, образованный складкой в мантии. Разница в размерах заднего и переднего концов этого органа определяет скорость струи, которую может производить организм. Скорость движения организма можно точно предсказать для данной массы и морфологии животного. Движение головоногих моллюсков обычно назад, так как вода вытесняется вперед через гипоном, но направление можно в некоторой степени контролировать, направляя его в разные стороны. Некоторые головоногие моллюски сопровождают это изгнание воды с похожим на выстрел хлопком, который, как считается, отпугивает потенциальных хищников.
Головоногие используют аналогичный метод движения, несмотря на то, что их увеличивающийся размер (по мере роста) меняет динамику движения. вода, в которой они оказались. Таким образом, их параларвы не слишком широко используют свои плавники (которые менее эффективны при низких числах Рейнольдса ) и в основном используют свои струи для продвижения вверх, тогда как большие взрослые головоногие моллюски имеют тенденцию плавать менее эффективно и больше полагаются на свои плавники.
 Nautilus belauensis вид спереди, показывающий раскрытие гипонома
Nautilus belauensis вид спереди, показывающий раскрытие гипонома Считается, что ранние головоногие моллюски производили струи, втягивая свое тело в свои раковины, как это делает сегодня наутилус. Наутилус также способен создавать струю волнообразными движениями своей воронки; этот более медленный поток воды больше подходит для извлечения кислорода из воды. Скорость струи у Наутилуса намного меньше, чем у колеоидов, но в ее производстве задействовано меньше мускулатуры и энергии. Реактивная тяга у головоногих в первую очередь определяется максимальным диаметром отверстия воронки (или, возможно, средним диаметром воронки) и диаметром мантийной полости. Изменения размера отверстия чаще всего используются при промежуточных скоростях. Достигаемая абсолютная скорость ограничена требованием головоногих моллюсков вдыхать воду для изгнания; этот прием ограничивает максимальную скорость восемью длинами тела в секунду, скорость, которую большинство головоногих может достичь после двух ударов воронкой. Вода наполняет полость, попадая не только через отверстия, но и через воронку. Кальмар может вытеснить до 94% жидкости внутри своей полости за одну струю. Чтобы приспособиться к быстрым изменениям водозабора и вытеснения, отверстия очень гибкие и могут изменять свой размер в двадцать раз; радиус воронки, наоборот, изменяется только примерно в 1,5 раза.
Некоторые виды осьминогов также могут ходить по морскому дну. Кальмары и каракатицы могут перемещаться на короткие расстояния в любом направлении за счет движения лоскута мышцы вокруг мантии.
Хотя большинство головоногих моллюсков плавают (т. Е. нейтрально плавучие или почти так; на самом деле большинство головоногих моллюсков примерно на 2–3% плотнее морской воды), они достигают этого разными способами. Некоторые, такие как Nautilus, позволяют газу диффундировать в зазор между мантией и оболочкой; другие позволяют более чистой воде сочиться из почек, вытесняя более плотную соленую воду из полости тела; другие, как некоторые рыбы, накапливают масла в печени; а у некоторых осьминогов студенистое тело с более легкими ионами хлора , замещающими сульфат в химическом составе тела.
Macrotritopus defilippi, или песок- жилого осьминога, имитирующего окраску и плавательные движения песчаной камбалы Bothus lunatus, чтобы избежать хищников. Осьминоги могли расплющивать свои тела и складывать руки назад, чтобы выглядеть так же, как камбалы, а также двигаться с той же скоростью и движениями.
Самки двух видов, Ocythoe tuberculata и Haliphron atlanticus, эволюционировали истинный плавательный пузырь.
 Поперечный разрез Spirula spirula, показывающий положение раковины внутри мантии
Поперечный разрез Spirula spirula, показывающий положение раковины внутри мантии 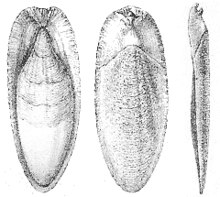 каракатицы из Sepia officinalis
каракатицы из Sepia officinalis  Gladius из Sepioteuthis lessoniana
Gladius из Sepioteuthis lessoniana Наутилусы - единственные сохранившиеся головоногие моллюски с истинной внешней оболочкой. Однако все раковины моллюсков образованы из эктодермы (внешнего слоя эмбриона); у каракатицы (Sepia spp.), например, инвагинация эктодермы формируется во время эмбрионального периода, в результате чего образуется раковина (каракатица ), которая находится внутри взрослого человека. То же самое относится и к хитиновому гладиусу кальмаров и осьминогов. Круглые осьминоги имеют дугообразные опоры хрящевых плавников, которые иногда называют «панцирем». рудимент »или« гладиус ». У Инциррина либо пара стержней, либо нет следов внутреннего панциря, а у некоторых кальмаров также нет гладиуса. Колеоиды в панцире не образуют кладу или даже парафилетическую группу. Оболочка спирулы начинается как органическая структура, а затем очень быстро минерализуется. «Потерянная» скорлупа может быть потеряна при резорбции карбонатного компонента кальция.
Самки рода осьминогов Argonauta выделяют специальный тонкий ящик для яиц, в котором они находятся, и это обычно считается «панцирем», хотя он не прикреплен к телу животного и имеет отдельное эволюционное происхождение.
Самая большая группа головоногих моллюсков, аммониты, вымерли, но их раковины очень распространены как окаменелости.
Отложения карбоната, приводящие к минерализованной раковине, по всей видимости, связано с кислотностью органической матрицы оболочки (см. оболочка моллюска ); Головоногие моллюски, образующие раковину, имеют кислый матрикс, тогда как гладиус кальмаров имеет основной матрикс. Основное устройство внешней стенки головоногих моллюсков: внешний (сферолитовый) призматический слой, ламинарный (перламутровый) слой и внутренний призматический слой. Толщина каждого слоя зависит от таксонов. У современных головоногих моллюсков карбонатом кальция является арагонит. Что касается других раковин моллюсков или скелетов кораллов, самые маленькие видимые единицы представляют собой неправильные округлые гранулы.

 Слева: гигантский кальмар, найденный в Логи-Бэй, Ньюфаундленд в 1873 году. Слева и справа видны два длинных щупальца для кормления.. Справа: Деталь щупальца Abraliopsis morisi
Слева: гигантский кальмар, найденный в Логи-Бэй, Ньюфаундленд в 1873 году. Слева и справа видны два длинных щупальца для кормления.. Справа: Деталь щупальца Abraliopsis morisi Головоногие, как следует из названия, имеют мускулистые придатки, выходящие из головы и окружающие рот. Они используются для кормления, передвижения и даже воспроизводства. В колеоидах их восемь или десять. У декапод, таких как каракатицы и кальмары, пять пар. Два более длинных, называемых щупальцами, активно участвуют в поимке добычи; они могут быстро удлиняться (всего за 15 миллисекунд). В гигантских кальмарах они могут достигать в длину 8 метров. Они могут оканчиваться расширенной булавой, покрытой присосками. Более короткие четыре пары называются руками и участвуют в удерживании захваченного организма и манипулировании им. У них тоже есть присоски на стороне, ближайшей ко рту; они помогают удержать добычу. У осьминогов есть только четыре пары покрытых присосками рук, как следует из названия, хотя аномалии развития могут изменять количество выраженных рук.
Щупальце состоит из толстого центрального нервного шнура (который должен быть толстым, чтобы позволить каждому присоска, которую нужно контролировать независимо), окруженная круговыми и радиальными мышцами. Поскольку объем щупальца остается постоянным, сокращение круговых мышц уменьшает радиус и позволяет быстро увеличивать длину. Как правило, удлинение на 70% достигается за счет уменьшения ширины на 23%. У более коротких рук такой возможности нет.
Размер щупальца зависит от размера ротовой полости; более крупные и сильные щупальца могут удерживать добычу, когда с нее получают небольшие укусы; с более многочисленными и меньшими щупальцами добыча проглатывается целиком, поэтому ротовая полость должна быть больше.
Внешне очищенные наутилиды (Nautilus и Allonautilus ) имеют порядка 90 пальцевидных придатков, называемых щупальцами, у которых отсутствуют присоски, но вместо этого они липкие и частично втягиваются.
 Двухчастный клюв гигантского кальмара, Architeuthis sp.
Двухчастный клюв гигантского кальмара, Architeuthis sp. У всех живых головоногих моллюсков клюв, состоящий из двух частей; у большинства из них имеется радула, хотя у большинства осьминогов она уменьшена, а у Spirula она вообще отсутствует. Они питаются, захватывая добычу щупальцами, втягивая ее в рот и кусая ее. У них есть смесь токсичных пищеварительных соков, некоторые из которых производятся симбиотическими водорослями, которые они выбрасывают из своих слюнных желез на захваченную добычу, которую они держат во рту. Эти соки отделяют плоть своей добычи от кости или панциря. На конце слюнной железы есть небольшой зуб, который можно воткнуть в организм, чтобы переварить его изнутри.
Сама пищеварительная железа довольно короткая. Он состоит из четырех элементов: пища проходит через зоб, желудок и слепую кишку, прежде чем попасть в кишечник. Большая часть пищеварения, а также всасывание питательных веществ происходит в пищеварительной железе, которую иногда называют печенью. Обмен питательными веществами и отходами между кишечником и пищеварительной железой осуществляется через пару соединений, соединяющих железу с местом соединения желудка и слепой кишки. Клетки пищеварительной железы непосредственно выделяют пигментированные экскреторные химические вещества в просвет кишечника, которые затем связываются со слизью, проходящей через задний проход в виде длинных темных нитей, выбрасываемых с помощью выдыхаемой воды из воронки. Головоногие, как правило, концентрируют проглоченные тяжелые металлы в тканях своего тела.
 Amphioctopus marginatus поедание краба
Amphioctopus marginatus поедание краба Радула головоногих моллюсков состоит из нескольких симметричных рядов до девяти зубов - тринадцать в ископаемых классы. У некоторых видов осьминогов этот орган уменьшен или даже рудиментарен и отсутствует у Spirula. Зубы могут быть гомодонтами (то есть подобными по форме в ряду), гетеродонтами (иначе) или гребневидными (гребенчатыми). Их высота, ширина и количество бугорков у разных видов различны. Рисунок зубов повторяется, но каждый ряд может не совпадать с предыдущим; у осьминога, например, последовательность повторяется каждые пять рядов.
Cephalopod radulae известны по ископаемым месторождениям, относящимся к ордовику. Обычно они сохраняются в камере тела головоногих, обычно в соединении с челюстями; но это не всегда так; многие радулы сохранились в различных местах в ручье Мейсон. Радулы обычно трудно обнаружить, даже если они сохранились в окаменелостях, поскольку порода должна выветриваться и трескаться именно так, чтобы обнажить их; например, радулы были обнаружены только у девяти из 43 родов аммонитов, и они еще реже встречаются в неаммоноидных формах: только три домезозойских вида обладают одним.
Большинство головоногих моллюсков обладают единственной парой крупных нефридий. Отфильтрованные азотистые отходы образуются в перикардиальной полости жаберного сердца, каждая из которых связана с нефридием узким каналом. Канал доставляет экскременты в почечный мешок, похожий на мочевой пузырь, а также поглощает лишнюю воду из фильтрата. Несколько отростков боковой полой вены выступают в почечный мешок, непрерывно раздуваясь и сдуваясь по мере биения жаберных сердец. Это действие помогает перекачивать секретируемые отходы в мешочки, чтобы они попали в полость мантии через поры.
Наутилус, что необычно, обладает четырьмя нефридиями, ни один из которых не связан с полостями перикарда.
Включение аммиака важно для образования раковины у наземных моллюсков и других немоллюсковых линий. Поскольку белок (т.е. мясо) является основным компонентом рациона головоногих, большие количества ионов аммония образуются как отходы. Основными органами, участвующими в выбросе этого избытка аммония, являются жабры. Скорость высвобождения самая низкая у головоногих моллюсков Nautilus и Sepia в результате того, что они использовали азот для наполнения раковины газом для увеличения плавучести. Другие головоногие моллюски используют аммоний аналогичным образом, накапливая ионы (как хлорид аммония ) для уменьшения общей плотности и увеличения плавучести.
 Самка Argonauta argo с яйцом и яйцами
Самка Argonauta argo с яйцом и яйцами  Деталь hectocotylus из Ocythoe tuberculata
Деталь hectocotylus из Ocythoe tuberculata  Расчлененный экземпляр самца Onykia ingens, показывает неэрегированный пенис (белая трубчатая структура, расположенная под большинством других органов)
Расчлененный экземпляр самца Onykia ingens, показывает неэрегированный пенис (белая трубчатая структура, расположенная под большинством других органов)  Образец того же вида, демонстрирующий удлинение пениса до 67 см в длину
Образец того же вида, демонстрирующий удлинение пениса до 67 см в длину Головоногие моллюски представляют собой разнообразную группу видов, но имеют общие черты жизненного цикла, например, они имеют быстрый темп роста и короткую продолжительность жизни. Стернс (1992) предположил, что для получения как можно большего числа жизнеспособных потомков нерест зависит от экологических факторов окружающей среды организма. Большинство головоногих моллюсков не заботятся о своем потомстве, за исключением, например, осьминога, который помогает этому организму повысить выживаемость их потомства. На жизненный цикл морских видов влияют различные условия окружающей среды. На развитие эмбриона головоногих моллюсков могут сильно влиять температура, насыщение кислородом, загрязнение, интенсивность света и соленость. Эти факторы важны для скорости эмбрионального развития и успеха вылупления эмбрионов. Доступность пищи также играет важную роль в репродуктивном цикле головоногих моллюсков. Ограничение пищи влияет на время нереста, а также на их функции и рост. Время нереста и нереста зависят от морских видов; это коррелирует с температурой, хотя головоногие моллюски на мелководье нерестятся в холодные месяцы, чтобы потомство вылупилось при более высоких температурах. Размножение может длиться от нескольких дней до месяца.
Половозрелые головоногие моллюски взрослого размера начинают нерест и размножаться. После передачи генетического материала следующему поколению взрослые головоногие моллюски умирают. Половое созревание у самцов и самок головоногих можно наблюдать изнутри по увеличению гонад и добавочных желез. Спаривание было бы плохим индикатором полового созревания самок; они могут получать сперму, когда она еще не полностью репродуктивно созрела, и хранить ее до тех пор, пока они не будут готовы к оплодотворению яйцеклеток. Самцы более агрессивны в соревновании перед спариванием в присутствии неполовозрелых самок, чем в соревновании за половозрелую самку. У большинства самцов головоногих моллюсков образуется гектокотиль, кончик руки, который способен переносить их сперматозоиды в полость камина самки. Хотя не все виды используют гектокотиль; например, взрослый наутилус выпускает спадикс. Признаком половой зрелости самок является развитие плечевых фотофоров для привлечения самок.
Головоногие моллюски не нерестовые производители. В процессе оплодотворения самки используют сперму, полученную от самца посредством внешнего оплодотворения. Внутреннее оплодотворение наблюдается только у осьминогов. Инициирование совокупления начинается, когда самец ловит самку и обхватывает ее рукой в положении «шея от мужчины к женщине» или в положении «рот к рту», в зависимости от вида. Затем самцы инициируют процесс оплодотворения, несколько раз сжимая мантию, чтобы высвободить сперматозоиды. Головоногие моллюски часто спариваются несколько раз, что заставляет самцов дольше спариваться с самками, у которых раньше было спаривание, что почти втрое увеличивает количество сокращений мантии. Чтобы обеспечить оплодотворение яйцеклеток, самки головоногих моллюсков выделяют привлекающий сперму пептид через студенистые слои яйца, чтобы направлять сперматозоиды. Самки головоногих моллюсков откладывают яйца в кладку; Каждое яйцо состоит из защитной оболочки, обеспечивающей безопасность развивающегося эмбриона при попадании в толщу воды. Репродуктивные стратегии различаются у разных видов головоногих. У гигантского тихоокеанского осьминога большие яйца откладывают в берлоге; Часто на их укладку уходит несколько дней. После того, как яйца высвобождаются и прикрепляются к защищенному субстрату, самки умирают, делая их полупородными. У некоторых видов головоногих кладки яиц прикрепляются к субстрату с помощью липкого липкого вещества. Эти яйца набухают перивителлиновой жидкостью (ПВФ), гипертонической жидкостью, предотвращающей преждевременное вылупление. Грозди оплодотворенных яиц обладают нейтральной плавучестью в зависимости от глубины, на которой они были отложены, но их также можно найти в таких субстратах, как песок, коралловая матрица или водоросли. Поскольку эти виды не заботятся о своем потомстве, капсулы яйца могут быть введены чернилами самкой, чтобы замаскировать эмбрионы от хищников.
Большинство головоногих моллюсков вступают в бой. при агрессивном сексе: белок в оболочке мужской капсулы стимулирует это поведение. Они также проявляют агрессию между мужчинами и мужчинами, при этом более крупные мужчины, как правило, выигрывают во взаимодействиях. Когда женщина находится рядом, самцы непрерывно атакуют друг друга и машут руками. Если ни один из самцов не отступает, руки вытягиваются назад, обнажая рот, после чего следует кусание кончиков рук. Во время соревнований самцов самцы также участвуют в технике, называемой смывом. Этот прием используется вторым самцом, пытающимся спариться с самкой. Промывка удаляет сперматофоры из ротовой полости, которые были помещены туда первым помощником, путем нагнетания воды в полость. Другое поведение, которое проявляют самцы, - это спаривание кроссовок или мимикрия - самцы меньшего размера подстраивают свое поведение под поведение самок, чтобы снизить агрессию. Используя эту технику, они могут оплодотворять яйца, в то время как более крупный самец отвлекается на другого самца. Во время этого процесса самцы кроссовок быстро вводят каплевидную сперму в семяприемник.
Выбор партнера наблюдается у видов каракатиц, у которых самки предпочитают одних самцов другим, хотя характеристики предпочтительного m эли неизвестны. Гипотеза утверждает, что самки отвергают самцов по обонятельным, а не по визуальным сигналам. Некоторые виды головоногих моллюсков принимают и хранят несколько мужских сперматофоров, что было идентифицировано с помощью ДНК-фингерпринта. Самки больше не восприимчивы к попыткам спаривания, когда держат яйца в руках. Самки могут хранить сперму в двух местах (1) в ротовой полости, куда недавно спарившиеся самцы помещают свои сперматофоры, и (2) во внутренних резервуарах для хранения спермы, где хранятся пакеты спермы от предыдущих самцов. Хранение сперматофора приводит к конкуренции сперматозоидов; в котором говорится, что самка контролирует, какой партнер оплодотворяет яйца. Чтобы уменьшить такого рода конкуренцию, самцы развивают агонистическое поведение, такое как охрана и смывание самки. Hapalochlaena lunulata, или осьминог с синими кольцами, легко спаривается как с самцами, так и с самками.
Наблюдается, что у многих морских организмов самки крупнее по сравнению с самцам у некоторых близкородственных видов. В некоторых линиях, таких как сплошной осьминог, самцы становятся все меньше и меньше по структуре, напоминая термин, карликовые самцы «карликовости» обычно встречаются при низкой плотности. Самец осьминога в одеяле - пример половой эволюционной карликовости; самки вырастают в 10 000-40 000 раз больше самцов, и соотношение полов между самцами и самками можно определить сразу после вылупления яиц.
 Яйца, отложенные самкой кальмаров
Яйца, отложенные самкой кальмаров Яйца головоногих моллюсков охватывают широкий диапазон размеров от 1 до 30 мм в диаметре. Оплодотворенная яйцеклетка сначала делится с образованием диска зародышевых клеток на одном полюсе, а желток остается на противоположном полюсе. Зародышевый диск растет, обволакивая и в конечном итоге поглощая желток, образуя зародыш. Щупальца и руки сначала появляются в задней части тела, где ступня была бы у других моллюсков, и только позже мигрируют к голове.
Воронка головоногих развивается на макушке головы, тогда как рот развивается на противоположной поверхности. Ранние эмбриологические стадии напоминают предковых брюхоногих и современных Monoplacophora.
Раковины развиваются из эктодермы как органический каркас, который впоследствии минерализуется. У Sepia, имеющей внутреннюю оболочку, эктодерма образует впячивание, поры которого закрываются до того, как этот органический каркас откладывается.
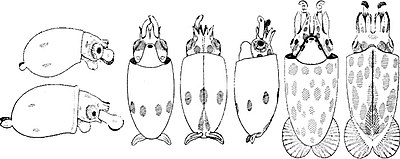 Chtenopteryx sicula paralarvae. Слева: Две очень молодые параларвы. Круглые щупальцевые булавы несут примерно 20 неправильно расположенных присосок. Два хроматофора присутствуют с каждой стороны мантии. В центре: вид спереди, сверху и сбоку более развитой параларвы. На мантии присутствует экваториальный круг из семи крупных желто-коричневых хроматофоров. Спереди при виде сверху видны расширенные лопасти гладиуса. Справа: вид очень развитой параларвы с вентральной и дорсальной стороны.
Chtenopteryx sicula paralarvae. Слева: Две очень молодые параларвы. Круглые щупальцевые булавы несут примерно 20 неправильно расположенных присосок. Два хроматофора присутствуют с каждой стороны мантии. В центре: вид спереди, сверху и сбоку более развитой параларвы. На мантии присутствует экваториальный круг из семи крупных желто-коричневых хроматофоров. Спереди при виде сверху видны расширенные лопасти гладиуса. Справа: вид очень развитой параларвы с вентральной и дорсальной стороны. 
 Слева: Незрелые экземпляры Chiroteuthis veranyi. В этой параларвальной форме, известной как стадия доратопсиса, перо длиннее мантии и шеи вместе взятых. Справа: зрелая Chiroteuthis veranyi. У этого вида одни из самых длинных щупалец по сравнению с размером любого известного головоногого моллюска.
Слева: Незрелые экземпляры Chiroteuthis veranyi. В этой параларвальной форме, известной как стадия доратопсиса, перо длиннее мантии и шеи вместе взятых. Справа: зрелая Chiroteuthis veranyi. У этого вида одни из самых длинных щупалец по сравнению с размером любого известного головоногого моллюска. Время до вылупления сильно варьируется; яйца меньшего размера в более теплой воде вылупляются быстрее всего, а новорожденные могут появиться уже через несколько дней. Более крупные яйца в более холодных водах могут развиваться в течение года до вылупления.
Процесс от нереста до вылупления у всех видов повторяется по схожей траектории, главной переменной является количество желтка, доступное молодняку, и время его появления. поглощается эмбрионом.
В отличие от большинства других моллюсков, головоногие моллюски не имеют морфологически отличной личиночной стадии. Вместо этого молодые особи известны как paralarvae. Они быстро учатся охотиться, используя встречи с добычей для уточнения своих стратегий.
Рост молодых особей обычно аллометрический, в то время как рост взрослых особей изометрический.
Традиционный взгляд на эволюцию головоногих моллюсков утверждает, что они произошли в позднем кембрии от моноплакофора -подобного предка с изогнутой сужающейся раковиной, который был тесно связан с брюхоногими ( улитки). В поддержку этой точки зрения использовалось сходство ранних головоногих моллюсков Plectronoceras с некоторыми брюхоногими моллюсками. Развитие сифункула позволило бы раковинам этих ранних форм стать наполненными газом (таким образом, плавучими), чтобы поддерживать их и удерживать раковины в вертикальном положении, пока животное ползло по полу, и разделяло настоящие головоногие моллюски от предполагаемых предков, таких как Knightoconus, у которых отсутствовал сифункул. Нейтральная или положительная плавучесть (то есть способность плавать) появилась бы позже, за ней следовало бы плавание в Plectronocerida и, в конечном итоге, реактивное движение у более производных головоногих.
Однако некоторые морфологические доказательства трудны. Чтобы примириться с этой точкой зрения, и повторное описание Nectocaris pteryx, у которого не было панциря и который, по-видимому, обладал реактивным движением наподобие "производных" головоногих, усложнило вопрос о порядке, в котором черты головоногих развились - при условии, что Nectocaris вообще является головоногим.
Ранние головоногие моллюски, вероятно, были хищниками на вершине пищевой цепи. После позднего кембрийского вымирания, которое привело к исчезновению многих аномалокарид, хищные ниши стали доступны для других животных. В ордовикский период примитивные головоногие моллюски претерпели импульсную диверсификацию, чтобы стать разнообразными и доминирующими в палеозое и мезозое морей.
В раннем палеозое их ареал был очень далеким. более ограниченный, чем сегодня; в основном они были приурочены к сублиторали неглубоких шельфов низких широт и обычно встречались в ассоциации с тромболитами. По мере развития ордовика постепенно принимался более пелагический облик. Глубоководные головоногие моллюски, хотя и редко, были обнаружены в нижнем ордовике, но только в водах высоких широт. В середине ордовика появились первые головоногие моллюски с перегородками, достаточно прочными, чтобы выдерживать давление, связанное с более глубокими водами, и они могли обитать на глубинах более 100–200 м. Направление свертывания раковины оказалось решающим для будущего успеха линий передачи; эндогастральное свертывание позволило бы достичь большого размера только с прямой оболочкой, тогда как экзогастральное свертывание - первоначально довольно редко - позволило развиться спиралям, знакомым по летописи окаменелостей, с соответствующими большими размерами и разнообразием. (Эндогастральный означает, что оболочка изогнута так, что брюшная или нижняя сторона вогнута в продольном направлении (брюшко внутрь); экзогастральная означает, что оболочка изогнута так, что брюшная сторона является продольно выпуклой (брюшко наружу), что позволяет воронке указывать назад под оболочку.)
 аммоноид с отсутствующей камерой тела, показывая поверхность перегородки (особенно справа) с ее волнистыми лопастями и седлами
аммоноид с отсутствующей камерой тела, показывая поверхность перегородки (особенно справа) с ее волнистыми лопастями и седлами Предки колеоидов (включая большинство современных головоногих) и предки современного наутилуса, разошелся во Флориде раннего ордовика, более 470 миллионов лет назад. Bactritida, силурийско-триасовая группа ортоконусов, широко считается парафилетической по отношению к колеоидам и аммоноидам, то есть последние группы возникли внутри Bactritida. Увеличение разнообразия колеоидов и аммоноидей наблюдается примерно в начале девонского периода и соответствует значительному увеличению разнообразия рыб. Это могло отражать происхождение двух производных групп.
В отличие от большинства современных головоногих моллюсков, большинство древних разновидностей имели защитные раковины. Эти раковины сначала были коническими, но позже превратились в изогнутые наутилоиды, которые наблюдаются у современных видов наутилусов. Считается, что конкурентное давление со стороны рыбы заставило оболочечные формы погрузиться в более глубокие воды, что оказало эволюционное давление на потерю панциря и привело к появлению современных колеоидов, изменение, которое привело к большим метаболическим затратам, связанным с потерей плавучести, но позволило их для повторного заселения мелководья. Однако некоторые из прямоболочечных наутилоидов превратились в белемнитов, из которых некоторые превратились в кальмаров и каракатиц. Утрата панциря могла также быть результатом эволюционного давления, направленного на повышение маневренности, что привело к появлению привычки, более напоминающей рыбу.
Были споры по поводу эмбриологического происхождения придатков головоногих. До середины двадцатого века гипотеза «Оружие как голова» была широко признана. Согласно этой теории, руки и щупальца головоногих моллюсков похожи на головные придатки брюхоногих моллюсков, предполагая, что они могут быть гомологичными структурами. Придатки головоногих моллюсков окружают рот, поэтому логично, что они могут быть получены из тканей головы эмбриона. Однако гипотеза «руки как ступня», предложенная Адольфом Наефом в 1928 году, получает все больше поддержки; например, картирование судеб зачатков конечностей в наутилусе с камерами указывает, что зачатки конечностей происходят из эмбриональных тканей «стопы».
Секвенирование полного генома головоногих моллюсков остается сложной задачей для исследователей из-за длины и повторяемости их ДНК. Первоначально предполагалось, что характеристики геномов головоногих являются результатом дупликаций всего генома. После полного секвенирования калифорнийского осьминога с двумя точками, в геноме были обнаружены сходные с другими морскими беспозвоночными паттерны со значительными добавками к геному, которые считаются уникальными для головоногих. Не было обнаружено никаких доказательств полной дупликации генома.
В геноме калифорнийского двухточечного осьминога есть существенные репликации двух семейств генов. Примечательно, что только ранее было известно, что расширенные генные семейства проявляют репликативное поведение у позвоночных. Первое семейство генов было идентифицировано как Протокадгерины, которые связаны с развитием нейронов. Протокадгерины функционируют как молекулы клеточной адгезии, необходимые для синаптической специфичности. Механизм репликации семейства генов протокадгерина у позвоночных приписывается сложному сплайсингу или разрезанию и вставке из локуса. После секвенирования калифорнийского двухточечного осьминога исследователи обнаружили, что семейство генов пророкадгерина у головоногих моллюсков расширилось в геноме из-за тандемной дупликации генов. Различные механизмы репликации генов протокадгерина указывают на независимую эволюцию экспансии генов протокадгерина у позвоночных и беспозвоночных. Анализ отдельных генов Cephalopod Protocadherin указывает на независимую эволюцию между видами Cephalopod. Вид прибрежных кальмаров Doryteuthis pealeii с расширенными семействами генов протокадгерина значительно отличается от видов калифорнийского двухточечного осьминога, что позволяет предположить, что экспансия гена не происходила до видообразования внутри головоногих моллюсков. Несмотря на разные механизмы экспансии генов, гены протокадгерина осьминога с двумя точками были больше похожи на позвоночных, чем на кальмаров, что указывает на механизм конвергентной эволюции. Второе семейство генов, известное как C2H2, представляет собой небольшие белки, которые функционируют как факторы транскрипции цинка. Считается, что C2H2 регулирует функции ДНК, РНК и белков в клетке.
Секвенированный геном калифорнийского двухточечного осьминога также показал значительное присутствие мобильных элементов, а также экспрессию транспозонов. Хотя роль мобильных элементов у морских позвоночных до сих пор относительно неизвестна, наблюдалась значительная экспрессия транспозонов в тканях нервной системы. В исследовании, проведенном на позвоночных, экспрессия транспозонов во время развития у Drosophila melanogaster активировала геномное разнообразие между нейронами. Это разнообразие связано с улучшением памяти и обучения у млекопитающих. Связь между транспозонами и повышенной способностью нейронов может дать представление об наблюдаемом интеллекте, памяти и функции головоногих.
Приблизительный консенсус существующей филогении головоногих моллюсков после Стругнелла и др. 2007, показан на кладограмме. Минерализованные таксоны выделены жирным шрифтом . Прикрепление клады, включая Сепию и Спирулу, неясно; любая из точек, отмеченных звездочкой, может представлять корень этого клады.
| Cephalopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Внутренняя филогения головоногих моллюсков трудно ограничить; многие молекулярные методы были приняты, но полученные результаты противоречивы. Nautilus, как правило, считается внешней группой, где Vampyroteuthis образует внешнюю группу по сравнению с другими кальмарами; однако в одном анализе наутилоиды, осьминоги и теутиды изображаются как политомия. Некоторые молекулярные филогении не выделяют минерализованные колеоиды (спирула, сепия и метасепия) в виде клады; однако другие действительно восстанавливают эту более скупую на вид кладу со спирулой в качестве клады. сестринская группа Sepia и Metasepia в кладе, которая, вероятно, разошлась до конца триаса.
Молекулярные оценки расхождения клад различаются. Согласно одной «статистически надежной» оценке наутилус отличался от осьминога на 415 ± 24 миллиона лет назад.
 Наутилус с камерой (Nautilus pompilius)
Наутилус с камерой (Nautilus pompilius)  Каракатица обыкновенная (Sepia officinalis)
Каракатица обыкновенная (Sepia officinalis)  Атлантический бобтейл (Sepiola atlantica)
Атлантический бобтейл (Sepiola atlantica)  Европейский кальмар (Loligo vulgaris)
Европейский кальмар (Loligo vulgaris)  Осьминог обыкновенный (Octopus vulgaris)
Осьминог обыкновенный (Octopus vulgaris) Представленная здесь классификация, для современных головоногих моллюсков следует в основном из Current Classification of Contemporary Cephalopoda (май 2001 г.), для ископаемых головоногих моллюсков - из Arkell et al. 1957 г., Тайхерт и Мур 1964 г., Тайхерт 1988 г. и др. Эти три подкласса являются традиционными, соответствующими трем отрядам головоногих, признанным Батером.
Класс Cephalopoda († указывает вымершие группы)
Другие классификации различаются, прежде всего, как связаны различные отряды декапод, и должны ли они быть отрядами или семьями.
Это более старая классификация, которая объединяет те, которые содержатся в частях K и L Трактата о палеонтологии беспозвоночных, которая составляет основу и в значительной степени сохраняется классификации, которые появились позже.
Наутилоиды в целом (Teichert and Moore, 1964) последовательность, как указано.
Палеозойские аммоноидеи (Miller, Furnish and Schindewolf, 1957)
Mesozoic Ammonoidea (Arkel et al., 1957)
Последующие изменения включают создание трех отрядов верхнего кембрия: Plectronocerida, Protactinocerida и Yanhecerida. ; разделение псевдортоцерид на псевдортоцериды и выделение ортоцератид как подкласса Orthoceratoidea.
Шевырев (2005) предложил разделение на восемь подклассов, в основном состоящих из более разнообразных и многочисленных ископаемых форм, хотя эту классификацию критиковали как произвольную.
 Gyronaedyceras eryx, наутилоид из девона в Висконсин
Gyronaedyceras eryx, наутилоид из девона в Висконсин  Различные виды аммонитов
Различные виды аммонитов  Голотип из семейства Ostenoteuthidae.
Голотип из семейства Ostenoteuthidae. Ископаемые класса белемнитов
Ископаемые класса белемнитов Cephalopoda
 Пиритизированное ископаемое Vampyronassa rhodanica, вампироморфид из нижнего келловея (166,1 миллионов лет назад)
Пиритизированное ископаемое Vampyronassa rhodanica, вампироморфид из нижнего келловея (166,1 миллионов лет назад) Другая недавняя система делит всех головоногих на две клады. Один включает наутилус и большинство ископаемых наутилоидов. Другая клада (Neocephalopoda или Angusteradulata) ближе к современным колеоидам и включает белемноидов, аммоноидей и многие семейства ортоцеридов. Есть также стволовая группа головоногих традиционных Ellesmerocerida, которые не принадлежат ни к одной кладе.
Колеоиды, несмотря на некоторые сомнения, судя по молекулярным данным, являются монофилетическими. 188>
 Ручка и Уот рисунок воображаемого колоссального осьминога, атакующего корабль, сделанный малакологом Пьером де Монфором, 1801
Ручка и Уот рисунок воображаемого колоссального осьминога, атакующего корабль, сделанный малакологом Пьером де Монфором, 1801 Древние мореплаватели знали о головоногих моллюсках, о чем свидетельствуют такие произведения искусства, как резьба по камню, найденная при археологических раскопках из бронзового века минойский Крит в Кносс (1900 - 1100 до н.э.). изображение рыбака, несущего осьминога. Ужасающе могущественная Горгона из греческой мифологии, возможно, была вдохновлена осьминогом или кальмаром, тело осьминога представляло отрубленную голову Медузы, а клюв - выступающий язык и клыки, а его щупальца - как змеи.
 Нашивка миссии NROL-39, изображающая Национальное разведывательное управление в виде осьминога с большим радиусом действия
Нашивка миссии NROL-39, изображающая Национальное разведывательное управление в виде осьминога с большим радиусом действия Кракен - легендарные морские чудовища гигантских размеров, которые, как говорят, обитают у берегов Норвегии и Гренландии, обычно изображаются в искусстве как гигантские головоногие моллюски, атакующие корабли. Линней включил его в первое издание своей книги Systema Naturae 1735 года. Гавайский миф о сотворении гласит, что нынешний космос - последний из серии, которая поэтапно возникла из руин предыдущей вселенной. В этом рассказе осьминог - единственный выживший из предыдущей, инопланетной вселенной. Аккорокамуи - это гигантский монстр с щупальцами из айну фолклора.
Сражение с осьминогом играет важную роль в Викторе Гюго Книга Travailleurs de la mer (Морские трудящиеся ), относящаяся к его пребыванию в изгнании на Гернси. Короткометражный фильм Яна Флеминга 1966 г. Сборник историй Octopussy и The Living Daylights, а также фильм 1983 Джеймс Бонд отчасти вдохновлены книгой Хьюго.
Японское эротическое искусство, сюнга, включает в себя укиё-э гравюры на дереве, такие как Кацусика Хокусай из 1814 года «Тако то ама» (Сон жены рыбака ), в котором ама дайвер сексуально переплетен с большим и малым осьминогами. Гравюра - предшественник эротики с щупальцами. Биолог П. З. Майерс отметил в своем научном блоге Pharyngula, что осьминоги появляются на «необычных» графических иллюстрациях, изображающих женщин, щупальца и обнаженную грудь.
Поскольку у них исходит множество рук. из общего центра, осьминог часто используется как символ мощной и манипулятивной организации, обычно отрицательно.
| Викивиды содержат информацию, относящуюся к Cephalopoda |
| В Wikibook Dichotomous Key есть страница по теме: Cephalopoda |
| Wikisource содержит текст Британской энциклопедии 1911 года статьи Cephalopoda. |
| Викискладе есть материалы по теме кому: . Cephalopoda. |





