
Войти
| Cartorhynchus. Временной диапазон: Spathian,. 248,41 Ma PreꞒ Ꞓ O S D C P T J K Pg N ↓ | |
|---|---|
 | |
| Трехмерная реконструкция черепа, вид снизу слева, обнажающая зубы и внутреннюю крышу черепа | |
| Научная классификация | |
| Домен: | Eukaryota |
| Царство: | Animalia |
| Тип: | Chordata |
| Класс: | Reptilia |
| Clade: | †Ichthyosauromorpha |
| Clade: | †Ichthyosauriformes |
| Clade: | †Nasorostra |
| Род: | †Cartorhynchus. Motani et al., 2014 |
| Виды: | †C. lenticarpus |
| Биномиальное название | |
| † Cartorhynchus lenticarpus . Motani et al., 2014 | |
Cartorhynchus (что означает «укороченная морда») - вымерший род ранних ichthyosauriform морские рептилии, которые жили в раннем триасе эпоху, около 248 миллионов лет назад. Род содержит единственный вид, Cartorhynchus lenticarpus, названный в 2014 году Риосуке Мотани и его коллегами из единственного почти полного скелета, найденного около Чаоху, провинция Аньхой, Китай. Вместе со своим близким родственником Sclerocormus, Cartorhynchus был частью диверсификации морских рептилий, которая возникла внезапно (примерно за миллион лет) во время спатского подэтапа, вскоре после разрушительного пермско-триасового вымирания, но впоследствии они были истреблены из-за вулканизма и изменений уровня моря в среднем триасе.
При длине около 40 сантиметров (16 дюймов) Cartorhynchus был небольшое животное с телом, напоминающим ящерицу, и коротким туловищем; вероятно, он плавал, как угорь, на медленных скоростях. Его конечности имели обширный хрящ и могли сгибаться, как ласты, что, возможно, позволяло ему ходить по суше. Самыми отличительными чертами Cartorhynchus были его короткая суженная морда и многочисленные ряды коренных -подобных зубов, которые росли на внутренней поверхности его челюстных костей. Эти зубы не были обнаружены до тех пор, пока образец не был подвергнут компьютерной томографии. Cartorhynchus, вероятно, охотился на беспозвоночных с твердым панцирем, используя всасывающее питание, хотя как именно он использовал свои направленные внутрь зубы, пока не известно. Это было одно из пяти независимых приобретений моляровидных зубов среди ихтиозавров.
 Стратиграфическая диаграмма формации Нанлингху в карьере Майяшан (место происхождения Sclerocormus отмечен красным)
Стратиграфическая диаграмма формации Нанлингху в карьере Майяшан (место происхождения Sclerocormus отмечен красным) В 2011 году единственный известный экземпляр Cartorhynchus был обнаружен в слое 633 на втором уровне карьера Маджашан недалеко от центра города Чаоху, провинция Аньхой, Китай ; пласты горных пород в этом карьере принадлежат к верхней пачке. Экземпляр состоит из почти полного скелета без только части хвоста и некоторых костей левой части задней части черепа. Сохранность образца, вероятно, была результатом того, что он был отложен в осадке правой стороной вниз, таким образом оставив левую сторону открытой для элементов. Он получил полевой номер MT-II, а позже номер образца AGB 6257 на.
В 2014 году этот образец был описан Риосуке Мотани и его коллегами в Nature как представляющий новый род и вид, Cartorhynchus lenticarpus. Они получили родовое имя Cartorhynchus от греческих слов kartos (καρτοσ, «сокращенный») и rinchios (ρηψνχηοσ, «морда»), а специфическое имя lenticarpus - от латинских слов lentus («гибкий») и запястье («запястье»). Оба названия относятся к анатомическим характеристикам, которые он имел бы при жизни. Образец считался беззубым, пока не был обнаружен изолированный зуб во время дальнейших попыток удалить камень между закрытыми челюстями. Поскольку образец был слишком хрупким, чтобы обнажить внутреннюю часть челюстей, Цзянь-Донг Хуанг, Мотани и другие коллеги впоследствии отсканировали и визуализировали образец в 3D с помощью микрокомпьютерной томографии (микро-КТ), выполненной в компании Yinghua Testing Company в Шанхае, Китай. В 2020 году результаты их последующей работы были опубликованы в Scientific Reports.
 Сравнение размеров
Сравнение размеров На момент своего открытия Cartorhynchus был самым маленьким известным представителем Ichthyosauriformes.. Сохранившийся образец имел длину 21,4 см (8 дюймов); Предполагая, что у него были пропорции хвоста, сопоставимые с пропорциями хвоста близких родственников, Мотани и его коллеги оценили полную длину тела в 40 сантиметров (1 фут 4 дюйма) и вес в 2 килограмма (4,4 фунта).
У Карторинха была необычно короткая и сжатая морда, которая занимала только половину длины черепа, и глубокая челюсть. Кончик морды был всего 6 миллиметров (0,2 дюйма) в ширину. В отличие от большинства рептилий, его носовая кость доходила до передней части морды. Из-за его также удлиненной предчелюстной кости его костные ноздри были расположены относительно далеко позади черепа, а его лобная кость также не имела расширения в его заднем внешнем углу. Все эти характеристики были общими с его близким родственником Sclerocormus. Однако, в отличие от последнего, лобная кость не вносила вклад в глазницу у Cartorhynchus; префронтальная и постфронтальная кости не соприкасались над глазницей; и расположение большого отверстия для шишковидной железы на крыше черепа было разным: оно было в месте контакта лобной и теменной костей у Sclerocormus, но только на теменных костях у Cartorhynchus. Cartorhynchus также имел характерно большие подъязычные кости.
 микро-КТ черепа, показывающие сужения (желтые стрелки) и поверхности износа (желтые скобки) на зубах
микро-КТ черепа, показывающие сужения (желтые стрелки) и поверхности износа (желтые скобки) на зубах Первоначально Мотани и его коллеги предположили, что Cartorhynchus был беззубым ; тем не менее, сканирование с помощью микро-КТ впоследствии показало наличие округлых, моляров -подобных зубов, которые выступали внутрь почти перпендикулярно длинной оси челюсти, что делало их невидимыми снаружи. Все зубы были либо полностью сплюснутыми, либо слабо заостренными, и многие зубы имели сужение между корнем и коронкой. В отличие от других ихтиозавров с коренными зубами, все коронки зубов были «раздутыми» в одинаковой степени. На верхней челюсти (кость верхней челюсти) и зубной части (кость нижней челюсти) зубы располагались в три ряда, причем самый внешний ряд имел самые большие зубы; у верхнечелюстных костей по семь, пять и, вероятно, по одному зубу, а у зубных костей по десять, семь и четыре зуба. Среди ихтиозавров только Cartorhynchus и Xinminosaurus имеют несколько рядов зубов. Расположение зубов означало, что самые передние нижние зубы не имели соответствующего верхнего зуба, а также что два зубных протеза, образующие нижнюю челюсть, не могли быть плотно слиты. Эта характеристика была бы характерна для его близких родственников, беззубых hupehsuchians.
 Реконструкция жизни
Реконструкция жизни Cartorhynchus, по-видимому, имел 5 шейных позвонков и 26 спинных позвонков. позвонки, всего 31 прекрестцовый позвонок (позвонок перед его крестцом или бедром). Наряду со Sclerocormus (с 34 пре-крестцовыми) и Chaohusaurus (с 36 пре-сакральными), Cartorhynchus попадает в типичный диапазон для наземных животных, в отличие от 40-80 пре-крестцов, распространенных среди более производный (специализированный) ихтиоптериги. В отличие от Sclerocormus, нервные отростки, выступающие из вершины позвонков у Cartorhynchus, были относительно узкими и наклонными, а не широкими и фланцевыми. Cartorhynchus также можно отличить по парапофизам, позвоночным отросткам, которые соединяются с ребрами; их передние края сливались с краями позвонков.
И Cartorhynchus, и Sclerocormus имели крепкие грудные клетки, которые были наиболее глубокими около плеча, с широкими, плоскими и толстостенными ребрами, что обычно наблюдается у ранних представителей вторично-водных рептилий. У Ichthyopterygia эти уплощенные ребра исчезли, за исключением Mollesaurus. На нижней стороне груди gastralia Cartorhynchus были тонкими и стержневидными, в отличие от уплощенной «корзины» Sclerocormus, но у обоих отсутствовала другая пара симметричных элементов по средней линии тела.
Конечности Карторинха были слабо окостеневшими (окостенели только три пальца руки) с широко расставленными костями, особенно между костями запястья (запястья) и пальцами., что свидетельствует о наличии обширного хряща в конечностях. Это сделало бы конечности подобными ластам. Ласты передних конечностей Cartorhynchus были изогнуты назад, при этом пальцы были наклонены на 50 ° относительно оси длинных костей (zeugopodium), в то время как задние конечности были изогнуты вперед. бедренная кость у Cartorhynchus была прямой и не расширена на нижнем конце. У Sclerocormus были похожие конечности, за исключением того, что они были лучше окостеневшими, а их сохранившаяся кривизна, возможно, не была естественной.
 Sclerocormus - ближайший родственник Cartorhynchus, причем две из них образуют Nasorostra
Sclerocormus - ближайший родственник Cartorhynchus, причем две из них образуют Nasorostra Отсутствие полных ископаемых останков привело к отсутствию ясности в отношении происхождения ихтиоптеригий, включая ихтиозавров. В течение многих лет считалось, что их окаменелости внезапно появились в среднем триасе с сильной водной адаптацией. Открытие Cartorhynchus и Sclerocormus частично восполнило этот пробел. Филогенетический анализ, проведенный Мотани и его коллегами, показал, что эти двое были тесно связаны друг с другом - образовывали кладу (группу), называемую Насоростра - и с ихтиоптеригией., к которой назоространс образовал сестринскую группу . Cartorhynchus и Sclerocormus были объединены их короткой мордой, удлиненными носами, глубокими челюстями, отсутствием расширения лобной части, наиболее глубокими грудными клетками около плеча и лопатками (лопатками), более широкими на нижнем конце, чем на верхнем конце..
Включение назоространса в филогенетический анализ также предоставило доказательства в поддержку hupehsuchians как близких родственников ихтиоптеригов. В 2014 году Мотани и его коллеги назвали кладу, образованную Nasorostra и Ichthyopterygia, Ichthyosauriformes, а кладу, образованную Ichthyosauriformes и Hupehsuchia, - Ichthyosauromorpha. Примечательно, что тесная связь между этими различными группами была восстановлена их анализами независимо от того, были ли из анализа удалены характеристики, связанные с водной адаптацией. Такие характеристики могли развиться гомоплазически (из конвергентной эволюции ) среди нескольких линий из-за сходного образа жизни, что может смещать филогенетический анализ, чтобы реконструировать их как гомологии (полученные из общая родословная). Устойчивость этих реконструированных отношений даже после удаления водных характеристик указывает на их надежность.
Ниже филогенетическое дерево из филогенетического анализа, опубликованного Хуангом, Мотани и коллегами в 2019 году, в описании Chaohusaurus brevifemoralis частично воспроизведен.
| Ichthyosauromorpha |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Появление ихтиозавроморфов было частью восстановления морских экосистем после разрушительного пермско-триасового вымирания. Обычно считалось, что морские экосистемы не восстановили свое полное разнообразие до 5-10 миллионов лет после массового вымирания, и что морские рептилии восстанавливались после исчезновения медленнее, чем другие линии. Однако открытие множества разнообразных фаун морских рептилий, обитавших в раннем триасе, включая Cartorhynchus, впоследствии показало, что это не так. В частности, оказывается, что ихтиозавриформы впервые появились на спатийском подэтапе оленекского этапа и быстро достигли высокого функционального разнообразия в первый миллион лет своей эволюции. Несмотря на относительно низкое видовое разнообразие, они занимали множество ниш, включая как демерсальные (обитающие на дне) виды, такие как hupehsuchians и nasorostrans, так и пелагические (открытые воды).) такие виды, как ихтиоптериги.
 Таксономическое разнообразие триасовых морских рептилий; обратите внимание на два пика разнообразия и узкое место между
Таксономическое разнообразие триасовых морских рептилий; обратите внимание на два пика разнообразия и узкое место между . Многие ихтиозавриформы из раннего и среднего триаса имели молярические зубы (показаны на филогенетическом дереве выше), включая Cartorhynchus; такие зубы указывают на диету, хотя бы частично основанную на животных с твердым панцирем. В 2020 году Хуанг и его коллеги выполнили реконструкцию предкового состояния зубов среди ихтиозавроморфов. Вероятностные методы предполагают, что округлые или плоские зубы, скорее всего, развивались независимо пять раз, в то время как методы, основанные на экономии, предполагают, что они развивались независимо от трех до пяти раз. Хуанг и его коллеги наблюдали, что развитие моляровидных зубов происходило независимо много раз у водных животных (включая несколько линий варан, мурен и спарид и цихлиды рыбы), и, таким образом, частота встречаемости среди ихтиозавриформ не является необычно высокой. Они также заметили, что у ихтиозавриформ раннего триаса обычно были маленькие округлые зубы; зубы ихтиозавриформ среднего триаса были более разнообразными по размеру и форме, что коррелирует с увеличением разнообразия беспозвоночных. Таким образом, они предположили, что диверсификация ихтиозавриформ была частично вызвана эволюцией добычи с твердым панцирем.
Однако hupehsuchians и nasorostrans в конечном итоге вымерли на границе между ранним и средним триасом, создав таксономическое узкое место ". На границе изменения уровня моря и вулканизм привели к плохо насыщенным кислородом океанам, что привело к возникновению характерной сигнатуры изотопа углерода от разлагающегося органического материала в пластах горных пород на границе. Разнообразие ихтиозавриформ после этого оборота не восстановилось: Sauropterygia и Saurosphargidae вызвали вторую волну диверсификации, длившуюся от трех до пяти миллионов лет.
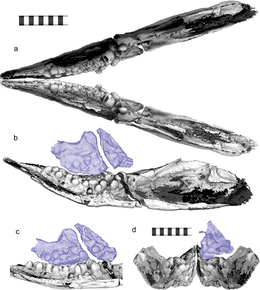 Трехмерная реконструкция зубных рядов и верхней челюсти в том виде, в каком они были бы сформулированы в жизни, в нескольких проекциях
Трехмерная реконструкция зубных рядов и верхней челюсти в том виде, в каком они были бы сформулированы в жизни, в нескольких проекциях Мотани и его коллеги предположили в 2014 году, что Cartorhynchus был всасывающим устройством, которое питалось концентрированием давление в его узкой морде. Это также подтверждается прочностью его подъязычной и поджаберной части (которые должны были закрепить язык), а также неправильным наблюдением беззубости. Впоследствии аналогичные выводы были сделаны для Sclerocormus. Shastasaurus и Shonisaurus ранее интерпретировались как ихтиозавры, питающиеся отсасыванием, но количественный анализ ихтиозавров триасового и юрского периодов, проведенный Мотани и его коллегами в 2013 году, показал, что ни у одного из них не было достаточно крепких гиобранхиальных костей или достаточно узкой морды, чтобы обеспечить всасывающее питание.
Обнаружение моляровидных зубов у Cartorhynchus привело Хуанга и его коллег к заключению в 2020 году, что Cartorhynchus был дурофагом, питаясь на добыче с твердым панцирем. Они отметили, что это не противоречит образу жизни с отсасыванием; некоторые спаридные рыбы являются одновременно дурофагами и питаются всасыванием. Однако они предположили, что Cartorhynchus был ограничен питанием мелкой добычей. Что касается горизонтальной ориентации зубов, они наблюдали поверхности износа, которые указывали на то, что стороны зубов перекрывались друг с другом, а не с коронками. Однако они отметили, что при высоких напряжениях раздавливающих укусов зубы являются структурно наиболее прочными на концах, а не на боковых сторонах. Они предположили, что челюсть могла быть искривлена во время консервации или что мягкие ткани, такие как коллаген, которые удерживали зубы при жизни, могли быть потеряны, хотя они признали, что ни одна из гипотез не может объяснить характер износа. Наконец, они отметили, что нижние зубы без соответствующих верхних зубов были необычными; на них нет следов износа, и нет никаких доказательств наличия мышечных механизмов, которые позволили бы использовать две челюсти друг против друга. Таким образом, они пришли к выводу, что эти зубы, вероятно, не использовались против других зубов.
 Cartorhynchus мог использовать свои ласты так же, как молодые морские черепахи
Cartorhynchus мог использовать свои ласты так же, как молодые морские черепахи У Cartorhynchus были плохо окостеневшие конечности несмотря на хорошо окостеневший череп и позвонки. Однако в 2014 году Мотани и его коллеги предположили, что он все еще взрослый, потому что многие ранне расходящиеся представители линий морских рептилий имеют плохо окостеневшие конечности из-за педоморфоза (сохранение незрелых черт в зрелом возрасте), хотя они и сделали. не полностью отвергать возможность того, что это был детеныш из-за существования только одного экземпляра.
В случае с Карторинчусом, Мотани и его коллеги предположили, что большие, похожие на ласты передние конечности позволили ему двигаться дальше. земли, что делает его амфибией. Обширный хрящ в лучезапястном суставе позволил бы плавнику сгибаться без локтя; У молодых морских черепах есть такие же хрящевые ласты, которые они используют для передвижения по суше. Хотя его плавник не был бы особенно сильным, Cartorhynchus был относительно легким, с соотношением массы тела к площади поверхности плавника меньше, чем у Chaohusaurus. Изогнутые ласты позволили бы держать их близко к телу, тем самым увеличивая их механическое преимущество. Мотани и его коллеги предположили, что другие черты Cartorhynchus также способствовали амфибийному образу жизни, в том числе короткое туловище и морда, а также утолщенные ребра (которые служили бы балластом, стабилизируя животное в прибрежной зоне. воды).
 Моделирование скорости потока; холодные цвета обозначают более медленное течение.
Моделирование скорости потока; холодные цвета обозначают более медленное течение. В исследовании 2019 года, проведенном Сусаной Гутарра и его коллегами, использовалось компьютерное моделирование для оценки затрат энергии на плавание у ихтиозавриформ. Ранние расходящиеся ихтиозавриформы с телом в форме ящерицы и удлиненными хвостами без лабиринтов, такие как Cartorhynchus, использовали бы угорьоподобное плавание, в то время как более поздние ихтиозавры с более глубокими, похожими на рыбу телами и хорошо выраженным хвостом двуустки использовали бы панцирно (похожее на макарель) плавание. Принято считать, что угловатое плавание менее эффективно, чем тигровое плавание. Действительно, Гутарра и его коллеги обнаружили, что энергетические затраты на плавание со скоростью 1 метр в секунду (3,3 фута / с) у Cartorhynchus были в 24-42 раза выше, чем у ихтиозавра Ophthalmosaurus (в зависимости от того, учитывается ли режим плавания.), а его коэффициент аэродинамического сопротивления был на 15% выше, чем у дельфина-афалины. Тем не менее, Cartorhynchus, вероятно, плавал с меньшей скоростью, что требовало меньшей эффективности, и преимущества плавания в форме каранжеи у более поздних, более крупных ихтиозавриформ были компенсированы увеличением размера тела.
 Chaohusaurus встречается в тех же слоях. как Cartorhynchus
Chaohusaurus встречается в тех же слоях. как Cartorhynchus Слой 633 карьера Majiashan, место, где был обнаружен Cartorhynchus, представляет собой слой серого глинистого (глинистого) известняка, расположенный в 13 метрах (43 фута) над основанием верхней пачки свиты Нанлинху. Он определяется сверху и снизу слоями желтоватого мергеля. С точки зрения биостратиграфии аммонита этот пласт относится к зоне. Оценки дат с высоким разрешением были произведены для отложений оленекского возраста, обнаженных в карьере Мацзяшан, на основе изотопных записей углерода-13 цикличности и спектрального гамма-излучения бревна (которые измеряют количество радиации в горных породах астрономического происхождения); В частности, слой 633 оценивается в возрасте 248,41 млн лет. В течение среднего триаса пласты Чаоху откладывались в океаническом бассейне относительно далеко от побережья, который был ограничен с юга более мелководными и карбонатными платформами, а с севера - континентальный склон и более глубокие бассейны.
Ихтиозавриформы Sclerocormus и Chaohusaurus обнаружены в карьере Маджашан вместе с Cartorhynchus; Sclerocormus известен из более молодого слоя 719 (248,16 млн лет), а Chaohusaurus встречается в обоих слоях. Зауроптеригий Majiashanosaurus известен из слоя 643. Разнообразие рыб в карьере Majiashan беднее, чем в других местах; наиболее распространенной рыбой является старейший известный представитель Perleidiformes, но также неизвестны широко распространенный вид Saurichthys и несколько неописанных рыб. Потенциальные беспозвоночные жертвы Cartorhynchus включают мелких аммонитов и двустворчатых моллюсков, а также тилакоцефалан членистоногих.