
Войти
| Членистоногие. Временной диапазон: 540–0 Ma До Ꞓ O S D C P T J K Pg N Кембрийский - недавний | |
|---|---|
 | |
| Вымершие и современные членистоногие. изображение сверху налево: † трилобит, † Stylonurus, скорпион, краб, сороконожка и бабочка | |
| Научная классификация | |
| Домен: | Эукариота |
| Царство: | Животное |
| Подцарство: | Эуметазоа |
| Клад: | ПараХоксозоа |
| Клад: | Bilateria |
| Clade: | Nephrozoa |
| (без рейтинга): | Protostomia |
| Superphylum: | Ecdysozoa |
| (без рейтинга): | Panarthropoda |
| (без рейтинга): | Tactopoda |
| Тип: | Arthropoda. von Siebold, 1848 |
| Subphyla, неразмещенные роды и классы | |
| |
| Разнообразие | |
| около 1,170,000 видов. | |
| Синонимы | |
Condylipoda Latreille, 1802 | |
An членистоногие (, от греческий ἄρθρον arthron, «сустав» и πούς pous, «стопа» (общ. ποδός)) - беспозвоночное животное, имеющее экзоскелет, сегментированное тело и парные сочлененные придатки. Членистоногие образуют тип Euarthropoda, который включает насекомых, паукообразных, многоножек и ракообразных. Первоначально предложенный термин Arthropoda относится к предлагаемой группировке Euarthropods и типу Onychophora.
Членистоногие характеризуются их сочлененными конечностями и кутикулой, сделанной из хитина., часто минерализованный с карбонатом кальция. Членистоногие в плане тела состоит из сегментов, каждый с парой придатков. Жесткая кутикула подавляет рост, поэтому членистоногие периодически заменяют ее линькой. Членистоногие двусторонне симметричны, и их тело имеет внешний скелет. У некоторых видов есть крылья.
Их универсальность позволила членистоногим стать наиболее богатыми видами членов всех экологических гильдий в большинстве сред. У них более миллиона описанных видов, что составляет более 80 процентов всех описанных видов живых животных, некоторые из которых, в отличие от большинства других животных, очень успешны в засушливой среде. Размер членистоногих варьируется от микроскопического ракообразного стиготантулюса до японского краба-паука.
Первичная внутренняя полость членистоногих - это гемоцель, в котором расположены его внутренние органы., и через которую циркулирует его гемолимфа - аналог крови ; у него открытая кровеносная система. Как и их внешность, внутренние органы членистоногих обычно состоят из повторяющихся сегментов. Их нервная система похожа на лестницу, с парными вентральными нервными тяжами, проходящими через все сегменты и образующими парные ганглии в каждом сегменте.. Их головы образованы путем слияния различного количества сегментов, а их мозг образован слиянием ганглиев этих сегментов и окружает пищевод. респираторная и экскреторная системы членистоногих различаются в зависимости как от среды обитания, так и от подтипа, к которому они принадлежат.
Их зрение зависит от различных комбинаций сложных глаз и пигментных ямок глазков : у большинства видов глазки могут определять только направление, с которого исходит свет, и составные глаза являются основным источником информации, но главные глаза пауков - это глазки, которые могут формировать изображения и, в некоторых случаях, могут поворачиваться, чтобы отслеживать добычу. У членистоногих также есть широкий спектр химических и механических датчиков, в основном на основе модифицированных ионы множества щетинок, известных как щетинок, которые выступают сквозь их кутикулу.
Способы размножения и развития членистоногих разнообразны; все наземные виды используют внутреннее оплодотворение, но это часто происходит путем непрямого переноса сперматозоидов через придаток или землю, а не путем прямой инъекции. Водные виды используют внутреннее или внешнее удобрение. Почти все членистоногие откладывают яйца, но скорпионы рождают живых детенышей после того, как яйца вылупляются внутри матери. Птенцы членистоногих варьируются от миниатюрных взрослых особей до личинок и гусениц, у которых отсутствуют суставные конечности,и в конечном итоге они претерпевают тотальный метаморфоз с образованием взрослой формы. Уровень материнского ухода за вылупившимися детенышами варьируется от полного отсутствия до длительного ухода со стороны скорпионов.
Эволюционное происхождение членистоногих восходит к кембрийскому периоду. Группа обычно рассматривается как монофилетическая, и многие анализы подтверждают размещение членистоногих с циклоневральными особями (или составляющими их кладами) в суперфилуме Ecdysozoa. В целом, однако, отношения базальных животных еще не решены. Точно так же до сих пор активно обсуждаются отношения между различными группами членистоногих.
Членистоногие вносят свой вклад в обеспечение человека пищей как непосредственно в качестве пищи, так и, что более важно, косвенно, как опылители сельскохозяйственных культур. Известно, что некоторые виды переносят тяжелые заболевания на людей, домашний скот и сельскохозяйственные культуры.
Слово «членистоногие» происходит от греческого ἄρθρον árthron, «сустав » и πούς pous (gen. podos (ποδός)), то есть «ступня» или «нога », что вместе означает «сочлененная нога». Обозначение «членистоногие» было придумано в 1848 году немецким физиологом и зоологом Карлом Теодором Эрнстом фон Зибольдом (1804–1885).
Членистоногие - это беспозвоночные с сегментированным телом и сочленёнными конечностями. экзоскелет или кутикула состоит из хитина, полимера глюкозамина. Кутикула многих ракообразных, клещей-жуков и многоножек (кроме щетинистых многоножек ) также биоминерализована с помощью карбоната кальция. Кальцификация эндостернита, внутренней структуры, используемой для прикрепления мышц, также встречается у некоторых opiliones.
Оценки количества видов членистоногих варьируются от 1 170 000 до 5-10 миллионов и составляют более 80 процентов всех известных видов животных. Количество видов остается трудно определить. Это происходит из-за допущений моделирования переписи, спроецированных на другие регионы с целью увеличения масштабов подсчетов в конкретных местах, применяемых ко всему миру. Поданным исследования 1992 года, только в Коста-Рике насчитывалось 500 000 видов животных и растений, из которых 365 000 - членистоногие.
Они являются важными членами морских, пресноводных, наземных и воздушных экосистем, и являются одной из двух основных групп животных, которые приспособились к жизни в засушливых условиях; другой - амниот, живыми членами которого являются рептилии, птицы и млекопитающие. Одна подгруппа членистоногих, насекомые, является самым богатым видами из всех экологических гильдий на суше и в пресноводных средах. Самые легкие насекомые весят менее 25 микрограммов (миллионных долей грамма), а самые тяжелые - более 70 граммов (2,5 унции). Некоторые живые ракообразные намного крупнее; например, лапы японского краба-паука могут достигать длины до 4 метров (13 футов), причем самым тяжелым из всех ныне живущих членистоногих является американский лобстер, превышающий 20 метров. кг (44 фунта).
 Голова _______________________ Грудь _______________________ Живот _______________________ Сегменты и тагматы членистоногого
Голова _______________________ Грудь _______________________ Живот _______________________ Сегменты и тагматы членистоногого  = Тело = Тазик (основание) = Жаберная ветвь // = Жаберные нити = Ножная ветвь Структура биравета отросток
= Тело = Тазик (основание) = Жаберная ветвь // = Жаберные нити = Ножная ветвь Структура биравета отросток эмбрионы всех членистоногих сегментированы, построены из серии повторяющихся модулей. Последний общий предок современных членистоногих, вероятно, состоял из ряда недифференцированных сегментов, каждый из которых имел пару придатков, которые функционировали как конечности. Однако всем известные живые и Ископаемые членистоногие сгруппировали сегменты в tagmata, в которых сегменты и их конечности специализированы по-разному.
Трехчастный внешний вид многих насекомых тел и двух- часть появления пауков является результатом этой группировки; фактически нет никаких внешних признаков сегментации у клещей. У членистоногих также есть два элемента тела, которые не являются частью этого последовательно повторяющегося паттерна сегментов: акрон спереди, перед ртом, и тельсон сзади, позади рта. анус. Глаза установлены на акроне.
Первоначально кажется, что каждый несущий придаток сегмент имел две отдельные пары придатков: верхнюю и нижнюю пару. Позже они сливаются в единую пару двояковыпуклых придатков, причем верхняя ветвь действует как жабра, а нижняя ветвь используется для передвижения. В некоторых сегментах всех известных членистоногих придатки были модифицированы, например, в форме жабр, частей рта, антенн для сбора информации или когтей для захвата; членистоногие «похожи на швейцарские армейские ножи, каждое из которых оснащено уникальным набором специальных инструментов». У многих членистоногих придатки исчезли из некоторых частей тела; особенно часто абдоминальные придаткиисчезают или сильно изменяются.
 Trilobitomorpha A L L L L L L Chelicerata x C P L L L L Ci Crustacea A A Mnd Mx Mx L L L L L Tracheata A x Mnd Mx Mx L L L L = acron = сегменты, влияющие на голову = сегменты тела x = потеряны в процессе развития = глаза = нефридии O = нефридии, утраченные в процессе развития A = Антенна L = ножка C = Chelicera P = Pedipalp Ci = Chilarium Mnd = Нижняя челюсть Mx = Maxilla Проблема с головой членистоногих
Trilobitomorpha A L L L L L L Chelicerata x C P L L L L Ci Crustacea A A Mnd Mx Mx L L L L L Tracheata A x Mnd Mx Mx L L L L = acron = сегменты, влияющие на голову = сегменты тела x = потеряны в процессе развития = глаза = нефридии O = нефридии, утраченные в процессе развития A = Антенна L = ножка C = Chelicera P = Pedipalp Ci = Chilarium Mnd = Нижняя челюсть Mx = Maxilla Проблема с головой членистоногих Наиболее заметная специализация сегментов находится в голове. Четыре основные группы членистоногих - Chelicerata (включает пауков и скорпионов ), Crustacea (креветок, лобстеры, крабы и т. Д.), Tracheata (членистоногие, которые дышат через каналы в свое тело; включает насекомых и многоножек ), а вымершие трилобиты - имеют головы, образованные из различных комбинаций сегментов, с отсутствующими придатками или по-разному специализированными. Кроме того, некоторые вымершие членистоногие, такие как Marrella, не принадлежат ни к одной из этих групп, поскольку их головы образованы их собственными особыми комбинациями сегментов и специализированных придатков.
Разработка эволюционного подхода Этапы, на которых могли появиться все эти различные комбинации, настолько сложны, что давно известны как «проблема головы членистоногих ». В 1960 году Р. Э. Снодграсс даже надеялся, что эта проблема не будет решена, поскольку он обнаружил, что попытки выработать решения были забавными.
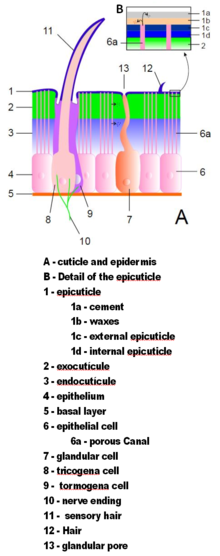 Иллюстрация идеализированного экзоскелета членистоногих.
Иллюстрация идеализированного экзоскелета членистоногих. Экзоскелеты членистоногих сделаны из кутикула, неклеточный материал, секретируемый эпидермисом. Их кутикула различается по деталям структуры, но обычно состоит из трех основных слоев: эпикутикулы, тонкого внешнего воскового покрытия, которое защищает другие слои от влаги и дает им некоторую защиту. ; экзокутикула, которая состоит из хитина и химически отвержденных белков ; и эндокутикула, которая состоит из хитина и незатвердевших белков. Экзокутикула и эндокутикула вместе известны как прокутикула. Каждый сегмент тела и часть конечности покрыты твердой кутикулой. Суставы между сегментами тела и между частями конечностей покрыты гибкой кутикулой.
Экзоскелеты большинства водных ракообразных биоминерализованы с помощью карбоната кальция извлекается из воды. Некоторые наземные ракообразные разработали способы хранения минерала, поскольку на суше они не могут полагаться на постоянное поступление растворенного карбоната кальция. Биоминерализация обычно затрагивает экзокутикулу и внешнюю часть эндокутикулы. Две недавние гипотезы об эволюции биоминерализации учленистоногих и других групп животных предполагают, что она обеспечивает более прочную защитную броню и что она позволяет животным становиться больше и сильнее, обеспечивая более жесткие скелеты; и в любом случае экзоскелет из минерально-органического композитного дешевле в изготовлении, чем полностью органический экзоскелет сопоставимой прочности.
Кутикула может иметь щетинок (щетинки) растет из особых клеток эпидермиса. Щетинки так же разнообразны по форме и функциям, как и придатки. Например, их часто используют в качестве датчиков для обнаружения воздушных или водных потоков или контакта с объектами; водные членистоногие используют перьевые -подобные щетинки для увеличения площади плавательных придатков и фильтрации частиц пищи из воды; водные насекомые, дышащие воздухом, используйте толстый войлок -подобный слой щетинок, чтобы задерживать воздух, увеличивая время пребывания под водой; тяжелые жесткие щетинки служат защитными шипами.
Хотя все членистоногие используют мышцы, прикрепленные к внутренней части экзоскелета, чтобы сгибать свои конечности, некоторые все еще используют гидравлическое давление, чтобы вытягивать их, система унаследована от их предков до членистоногих; например, все пауки гидравлически вытягивают ноги и могут создавать давление, в восемь раз превышающее их уровень покоя.
 Цикада вылезает из своего экзоскелета, будучи прикрепленным к дереву
Цикада вылезает из своего экзоскелета, будучи прикрепленным к дереву Экзоскелет не может растягиваются и тем самым ограничивают рост. Поэтому членистоногие заменяют свои экзоскелеты, подвергаясь экдизу (линьке) или сбрасывая старый экзоскелет после выращивания нового, еще не затвердевшего. Циклы линьки проходят почти непрерывно, пока членистоногие не достигнет полного размера.
Стадии развития между каждой линькой (шелушение) до достижения половой зрелости называются возрастом. Различия между возрастами часто можно увидеть в изменении пропорций тела, цвета, рисунка, изменении количества сегментов тела или ширины головы. После линьки, т. Е. Сбрасывания экзоскелета, молодые членистоногие продолжают свой жизненный цикл до тех пор, пока они не окукливаются или не линяют снова.
В начальной фазе линьки животное перестает питаться, и его эпидермис выделяет линяющую жидкость, смесь ферментов, которая переваривает эндокутикулу и, таким образом, отделяет старую кутикулу.. Эта фаза начинается, когда эпидермис секретирует новую эпикутикулу, чтобы защитить ее от ферментов, и эпидермис секретирует новую экзокутикулу, в то время как старая кутикула отделяется. Когда эта стадия завершается, животное набухает, поглощая большое количество воды или воздуха, и это заставляет старую кутикулу расщепляться по заранее определенным слабостям там, где старая экзокутикула была самой тонкой. Обычно животному требуется несколько минут, чтобы вырваться из старой кутикулы. На этом этапе новаяэндокутикула становится морщинистой и настолько мягкой, что животное не может поддерживать себя и ему очень трудно двигаться, а новая эндокутикула еще не сформировалась. Животное продолжает накачивать себя, чтобы максимально растянуть новую кутикулу, затем новая экзокутикула затвердевает и удаляет лишний воздух или воду. К концу этой фазы формируется новая эндокутикула. Многие членистоногие затем поедают выброшенную кутикулу, чтобы восстановить ее материалы.
Поскольку членистоногие не защищены и почти обездвижены, пока новая кутикула не затвердеет, они подвергаются опасности как попасть в старую кутикулу, так и подвергнуться нападению хищников.. Линька может быть причиной от 80 до 90% всех смертей членистоногих.
 = сердце = кишечник = мозг / ганглии 0 = глаз Основная структура тела членистоногих
= сердце = кишечник = мозг / ганглии 0 = глаз Основная структура тела членистоногих Тела членистоногих также сегментированы изнутри, а нервная, мышечная, кровеносная и выделительная системы имеют повторяющиеся компоненты. Членистоногие происходят от линии животных, у которых есть целом, выстланная мембраной полость между кишечником и стенкой тела, в которой находятся внутренние органы. Сильные сегментированные конечности членистоногих исключают необходимость в одной из основных наследственных функций целома, например, в гидростатическом скелете, мышцы которого сжимаются, чтобы изменить форму животного и, таким образом, дать ему возможность двигаться. Следовательно, целом у членистоногих сокращается до небольших участков вокруг репродуктивной и выделительной систем. Его место в основном занимает hemocoel, полость, которая проходит по большей части тела и через которую проходит кровь.
Членистоногие имеют открытую кровеносную систему, хотя у большинства из них есть несколько коротких, открытых артерий. У хелицератов и ракообразных кровь переносит кислород к тканям, тогда как гексаподы используют отдельную систему трахеи. Многие ракообразные, но мало хелицератов и трахеатов, используют респираторные пигменты для облегчения транспорта кислорода. Наиболее распространенным респираторным пигментом у членистоногих является гемоцианин на основе меди ; это используется многими ракообразными и некоторыми многоножками. Некоторые ракообразные и насекомые используют железосодержащий гемоглобин, респираторный пигмент, используемый позвоночными. Как и у других беспозвоночных, дыхательные пигменты тех членистоногих, у которых они есть, обычно растворены в крови и редко заключены в тельца, как у позвоночных.
Сердце обычно представляет собой мышечную трубку. который проходит прямо под спиной и на большей части длины гемоэля. Он сокращается в виде ряби, бегущей сзади вперед, выталкивая кровь вперед. Участки, не сдавливаемые сердечной мышцей,расширяются либо эластичными связками, либо небольшими мышцами, в любом случае соединяющими сердце со стенкой тела. По сердцу запустить серию парных устьев, невозвратных клапанов, которые позволяют крови попадать в сердце, но не позволяют ей выйти до того, как она достигнет передней части.
У членистоногих есть самые разные дыхательные системы. Маленькие виды часто не имеют его, так как их высокое отношение площади поверхности к объему обеспечивает простую диффузию через поверхность тела для обеспечения достаточного количества кислорода. У ракообразных обычно есть жабры, представляющие собой видоизмененные придатки. У многих паукообразных книжные легкие. Трахеи, системы разветвляющихся туннелей, которые выходят из отверстий в стенках тела, доставляют кислород непосредственно к отдельным клеткам многих насекомых, многоножек и паукообразных.
У живых членистоногих есть спаренные основные нервные тяжи проходят вдоль их тел ниже кишечника, и в каждом сегменте связки образуют пару ганглиев, от которых сенсорные и моторные нервы идут к другим частям сегмента.. Хотя пары ганглиев в каждом сегменте часто кажутся физически сросшимися, они связаны спайками (относительно большими пучками нервов), которые придают нервной системе членистоногих характерный "лестничный" вид. Мозг находится в голове, окружает пищевод и находится в основном над ним. Он состоит из сросшихся ганглиев акрона и одного или двух передних сегментов, образующих голову - всего три пары ганглиев у большинства членистоногих, но только две у хелицератов, которые не имеют усиков или связанных с ними ганглиев.. Ганглии других сегментов головы часто находятся рядом с мозгом и функционируют как его часть. У насекомых эти другие головные ганглии объединяются в пару субэзофагеальных ганглиев, расположенных под и позади пищевода. Пауки продвигают этот процесс еще дальше, поскольку все сегментарные ганглии включены в субэзофагеальные ганглии, которые занимают большую часть пространства в головогруди (передний «суперсегмент»).
Есть два разных типа выделительной системы членистоногих. У водных членистоногих конечным продуктом биохимических реакций, метаболизирующих азот, является аммиак, который настолько токсичен, что его нужно разбавлять как можно больше вода. Затем аммиак удаляется через любую проницаемую мембрану, в основном через жабры. Все ракообразные используют эту систему, и ее высокое потребление воды может быть причиной относительной неуспешности ракообразных как наземных животных. Различные группы наземных членистоногих независимо друг от друга разработали разные системы: конечным продуктом азотистого обмена является мочевая кислота, которая может выделяться в виде сухого вещества; система мальпигиевых канальцев фильтрует мочевую кислоту и другие азотистые отходыиз крови в гемоцеле и сбрасывает эти материалы в заднюю кишку, из которой они выводятся в виде фекалий. У большинства водных членистоногих и некоторых наземных также есть органы, называемые нефридии («маленькие почки »), которые выделяют другие отходы для выделения в виде мочи.
Жесткие кутикулы членистоногих блокируют информацию о внешнем мире, за исключением того, что через них проникают многие датчики или соединения от датчиков к нервной системе. Фактически, членистоногие превратили свою кутикулу в сложный массив сенсоров. Различные сенсорные датчики, в основном щетинки, реагируют на разные уровни силы, от сильного контакта до очень слабых воздушных потоков. Химические сенсоры обеспечивают эквиваленты вкуса и запаха, часто с помощью щетинок. Датчики давления часто имеют форму мембран, которые функционируют как барабанные перепонки, но подключены непосредственно к нервам, а не к слуховым косточкам. антенны большинства гексаподов включают пакеты датчиков, которые контролируют влажность, влажность и температуру.
 Голова осы с тремя глазки (в центре) и сложные глаза слева и справа
Голова осы с тремя глазки (в центре) и сложные глаза слева и справа Большинство членистоногих имеют сложные зрительные системы, которые включают один или несколько, обычно оба из сложных глаз и пигментных чашек глазков ( "глазки"). В большинстве случаев глазки способны определять направление, откуда исходит свет, только по тени, отбрасываемой стенками чашки. Однако главные глаза пауков представляют собой глазки пигментных чашечек, которые способны формировать изображения, а глаза прыгающих пауков могут вращаться, чтобы выслеживать добычу.
Составные глаза состоят из пятнадцати или нескольких тысяч независимых омматидий, столбцов, которые обычно шестиугольные в поперечном сечении. Каждый омматидий представляет собой независимый датчик со своими собственными светочувствительными клетками и часто со своей собственной линзой и роговицей. Составные глаза имеют широкое поле зрения и могут обнаруживать быстрое движение и, в некоторых случаях, поляризацию света. С другой стороны, относительно большой размер омматидиев делает изображения довольно грубыми, а сложные глаза более близорукие, чем у птиц. и млекопитающие - хотя это не является серьезным недостатком, поскольку объекты и события в пределах 20 сантиметров (7,9 дюйма) являются наиболее важными для большинства членистоногих. Некоторые членистоногие обладают цветовым зрением, а у некоторых насекомых было детально изучено; Например, омматидии пчел содержат рецепторы как для зеленого, так и для ультрафиолетового.
. Большинству членистоногих не хватает баланса и датчиков ускорения, и они полагаются на свои глаза, чтобы указать им, какой путь вверх. Самовосстанавливающееся поведение тараканов срабатывает, когда датчики давления на нижней стороне ступней не сообщают о давлении. Однако у многих малакостраканов ракообразных есть статоцисты, которые предоставляют такую же информацию, что и датчики баланса и движения позвоночного внутреннего уха.
проприорецепторы членистоногих, датчики, которые регистрируют силу, прилагаемую мускулами, и степень изгиба тела и суставов, хорошо изучены. Однако мало что известно о том, какие внутренние сенсоры могут иметь членистоногие.
 Самка Compsobuthus werneri с молодыми (белыми)
Самка Compsobuthus werneri с молодыми (белыми) Несколько членистоногих, например ракушки, гермафродиты, то есть каждый может иметь органы обоих полов. Однако особи большинства видов остаются одного пола на всю жизнь. Некоторые виды насекомых и ракообразных могут воспроизводиться посредством партеногенеза, особенно если условия благоприятствуют «популяционному взрыву». Однако большинство членистоногих полагаются на половое размножение, а партеногенетические виды часто возвращаются к половому размножению, когда условия становятся менее благоприятными. Водные членистоногие могут размножаться путем внешнего оплодотворения, как, например, лягушки. сделать, или посредством внутреннего оплодотворения, где яйцеклетки остаются в теле женщины, а сперматозоиды должны каким-то образом быть вставлены. Все известные наземные членистоногие используют внутреннее оплодотворение. Opiliones (уборщики), многоножки и некоторые ракообразные используют модифицированные придатки, такие как гононогие или пенисы, для передачи спермы непосредственно самке.. Однако большинство самцов наземных членистоногих производят сперматофоры, водонепроницаемые пакеты сперматозоидов, которые самки принимают в свои тела. Некоторые такие виды полагаются на самок, чтобы найти сперматофоры, которые уже были отложены на земле, но в большинстве случаев самцы откладывают сперматофоры только тогда, когда сложные ритуалы ухаживания кажутся вероятными.
 Личинка науплиуса a креветка-пенеид
Личинка науплиуса a креветка-пенеид Большинство членистоногих откладывают яйца, но скорпионы яйцекладущие : они производят живых детенышей после того, как яйца вылупились внутри матери, и отличаются длительной материнской заботой. Новорожденные членистоногие имеют разнообразные формы, и одни только насекомые охватывают множество крайностей. Некоторые вылупляются как явно миниатюрные взрослые особи (прямое развитие), а в некоторых случаях, например, серебрянка, вылупившиеся детеныши не питаются и могут быть беспомощными до тех пор, пока не начнется первая линька. Многие насекомые вылупляются в виде личинок или гусениц, у которых нет сегментированных конечностей или закаленной кутикулы, и превращаются во взрослыеформы, вступая в неактивную фазу, в которой ткани личинок разрушаются и восстанавливаются. - используются для построения тела взрослой особи. Личинки стрекозы имеют типичную кутикулу и суставные конечности членистоногих, но являются нелетающими водными дышащими животными с выдвижными челюстями. Ракообразные обычно вылупляются в виде крошечных личинок науплиусов, у которых всего три сегмента и пары придатков.
Последний общий предок всех членистоногих реконструирован как модульный организм, каждый модуль которого покрыт своим собственным склеритом (броневой пластиной) и несет пару бираманных конечностей. Однако вопрос о том, была ли предковая конечность одноранговой или двуручной, далеко не окончательный спор. У этого ур-членистоногого был вентральный рот, преоральные антенны и дорсальные глаза на передней части тела. Предполагалось, что это был недискриминационный донный питатель, перерабатывающий любой осадок, попавшийся на его пути в пищу, но находки окаменелостей намекают на то, что последний общий предок обоих членистоногих и приапулид разделял одно и то же специализированный ротовой аппарат; круглый рот с кольцами зубов, использовавшийся для поимки добычи и, следовательно, был плотоядным.
 Маррелла, одно из загадочных членистоногих из сланцев Берджесс
Маррелла, одно из загадочных членистоногих из сланцев Берджесс . предположил, что эдиакарские животные Парванкорина и Сприггина, появившиеся примерно в 555 миллионах лет назад, были членистоногими. Небольшие членистоногие с раковинами, напоминающими двустворчатые моллюски, были обнаружены в окаменелостях раннего кембрия, датируемых 541-539 миллионами лет назад в Китае и Австралии. Самые ранние окаменелости кембрийских трилобитов имеют возраст около 530 миллионов лет, но этот класс уже был довольно разнообразным и всемирным, предполагая, что они были здесь довольно давно. Повторное исследование в 1970-х годах окаменелостей сланца Берджесс примерно 505 миллионов лет назад выявило множество членистоногих, некоторые из которых не могли быть отнесены ни к одной из хорошо известных групп, и таким образом усилили дискуссию о кембрийском взрыве. Ископаемые останки Марреллы из сланцев Берджесс предоставили самые ранние четкие доказательства линьки.
Самые ранние ископаемые ракообразные датируются примерно 511 миллионами лет назад в Кембрийский и ископаемые креветки примерно 500 миллионов лет назад, очевидно, образовали сплоченную процессию по морскому дну. Окаменелости ракообразных распространены начиная с ордовика периода и далее. Они остались почти полностью водными, возможно потому, что у них никогда не развивались выделительные системы, сохраняющие воду.
Членистоногие представляют собой самые ранние идентифицируемые окаменелости наземных животных, примерно от 419 миллионов. лет назад, в позднем силурийском периоде, и земные следы примерно 450 миллионов лет назад, по-видимому, были сделаны членистоногими. Членистоногие были хорошо предварительно адаптированы для колонизации земли, потому что их существующие сочлененные экзоскелеты обеспечивали защиту от высыхания, поддержку от силы тяжести и средства передвижения, которые не зависели от воды. Примерно в то же время водные скорпионоподобные эвриптериды стали самыми крупными из когда-либо существовавших членистоногих, некоторые достигали 2,5 метра (8,2 фута).
Самый старый из известных паукообразных это тригонотарбид jerami, примерно с 420 миллионов лет назад в силурийский период. Attercopus fimbriunguis, с 386 миллион лет назад, в девонский период, здесь были самые ранние из известных шелковых кранов, но отсутствие фильер означает, что это не был один из настоящих пауков, которые впервые появились в конце карбона более 299 миллионов лет назад. юрский и меловой периоды дают большое количество ископаемых пауков, включая представителей многих современных семейств. Окаменелости водных скорпионов с жабрами появляются в силурийском и девонском периодах и являются самыми ранними останками дышащего воздухом скорпиона с книжными легкими датируется ранним каменноугольным периодом.
Самая старая окончательная окаменелость насекомых - это девон Rhyniognatha hirsti, датируемая 396-407 миллион лет назад, но его челюсти относятся к типу, встречающемуся только у крылатых насекомых, что позволяет предположить, что самые ранние насекомые появились в силурийский период. Mazon Creek lagerstätten из позднего карбона, около 300 миллионов лет назад, включает около 200 видов, некоторые из которых являются гигантскими по современным меркам, и указывают на то, что насекомые заселили свой основной современный экологические ниши как травоядные, детритофаги и насекомоядные. Социальные термиты и муравьи впервые появляются в раннем меловом периоде, а высокоразвитые социальные пчелы были обнаружены в позднемеловых породах, но не стали многочисленными до середины Кайнозой.
 бархатный червь (Onychophora) тесно связан с членистоногими
бархатный червь (Onychophora) тесно связан с членистоногими С 1952 по 1977 год зоолог Сидни Мантон и другие утверждали, что членистоногие полифилетичны, иными словами, у них нет общего предка, который сам был членистоногим. Вместо этого они предположили, что три отдельные группы «членистоногих» произошли отдельно от общих червеобразных предков: хелицератов, включая пауков и скорпионов ; ракообразные; и унирамия, состоящая из онихофора, многоножек и гексапода. Эти аргументы обычно игнорировались трилобитами, так как эволюционные отношения этого класса были неясны. Сторонники полифилии утверждали следующее: что сходство между этими группами является результатом конвергентной эволюции, как естественное следствие наличия жестких сегментированных экзоскелетов ; что все три группы используют разные химические средства для укрепления кутикулы; что были существенные различия в строении их сложных глаз; что трудно понять, как такие разные конфигурации сегментов и придатков головы могли развиться от одного и того же предка; и что ракообразные имеют двуветвистые конечности с отдельными жаберными и ножными ветвями, в то время как две другие группы имеют однообразные конечности, в которых одна ветвь служит ногой.
| ||||||||||||||||||||||||||||
Дальнейший анализ и открытия в 1990-е годы изменили эту точку зрения и привели к признанию того, что членистоногие монофилетичны, другими словами, у них действительно общий предок, который сам был членистоногим. Например, анализ Грэма Бадда Керигмачелы в 1993 году и Опабинии в 1996 году убедил его, что эти животные были похожи на онихофораны и различные ранние Кембрийский «лобопод », и он представил «эволюционное генеалогическое древо», которое показало их как «тетушек» и «кузенов» всех членистоногих. Эти изменения сделали неясным объем термина «членистоногие», и Клаус Нильсен предложил обозначить более широкую группу «Panarthropoda » («все членистоногие»), а животных с суставными конечностями и твердой кутикулой - называться «Euarthropoda» («настоящие членистоногие»).
Противоположная точка зрения была представлена в 2003 году, когда Ян Бергстрем и Сянь-Гуан Хоу утверждали, что если членистоногие были «сестринской группой» для любого из аномалокарид, они, должно быть, утратили, а затем заново развили черты, которые были хорошо развиты у аномалокарид. Самые ранние известные членистоногие ели грязь, чтобы извлекать из нее частицы пищи, и обладали переменным количеством сегментов с неспециализированными придатками, которые функционировали как жабры, так и ноги. Аномалокариды были, по стандартам того времени, огромными и сложными хищниками со специализированнымиртами и хватательными придатками, фиксированным количеством сегментов, некоторые из которых были специализированными, хвостовыми плавниками и жабрами, которые сильно отличались от таковых у членистоногих. Это рассуждение подразумевает, что Parapeytoia, у которого есть ноги и рот, направленный назад, как у самых ранних членистоногих, является более вероятным ближайшим родственником членистоногих, чем Anomalocaris. В 2006 году они предположили, что членистоногие более тесно связаны с лобоподами и тихоходками, чем с аномалокаридами. В 2014 году исследования показали, что тихоходки более тесно связаны с членистоногими, чем с бархатными червями.
| Протостомы |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Выше «родословной», Annelida традиционно считались ближайшими родственниками панартропод, поскольку обе группы имели сегментированные тела, и комбинация этих групп была обозначена как Articulata. Существовали конкурирующие предположения, что членистоногие были тесно связаны между собой к другим группам, таким как нематоды, приапулиды и тихоходки, но они остались взглядами меньшинства, потому что было трудно детально определить отношения между этими группами.
В 1990-х годах молекулярно-филогенетический анализ последовательностей ДНК позволил получить согласованную схему, показывающую членистоногих как членов супрофилума, обозначенного как Ecdysozoa («животные та линька »), который содержал нематод, приапулид и тихоходок, но исключал кольчатых червей. Это было подтверждено исследованиями анатомии и развития этих животных, которые показали, что многие из особенностей, которые поддерживали гипотезу Articulata, показали существенные различия между кольчатыми червями и самыми ранними панартроподами в деталях, а некоторые вообще не присутствовали у членистоногих. Эта гипотеза группирует кольчатых червей с моллюсками и брахиопод в другой супертип, Lophotrochozoa.
Если гипотеза Ecdysozoa верна, то сегментирование членистоногих и кольчатых червей либо эволюционировало конвергентно, либо были унаследованы от гораздо более древнего предка и впоследствии утеряны в нескольких других линиях, таких как представители Ecdysozoa, не являющиеся членистоногими.
Членистоногие принадлежат к типу Euarthropoda. Тип иногда называют Arthropoda, но строгоэтот термин обозначает кладу (предполагаемую - см. Tactopoda ), которая также включает филум Onychophora.
Euarthropoda, как правило, подразделяется на пять подтип, один из которых вымер:
Помимо этих основных групп, существует также ряд окаменелостей формы, в основном из раннего кембрия, которые трудно определить либо из-за отсутствия очевидной близости к какой-либо из основных групп, либо из-за явной близости к некоторым из них. Marrella был первым, кто был признан значительно отличным от хорошо известных групп.
Филогения основных современных групп членистоногих была областью значительных интерес и спор. Недавние исследования убедительно свидетельствуют о том, что Crustacea, как традиционно определяется, парафилетическим, с Hexapoda, развившейся внутри него, так что Crustacea и Hexapoda образуют кладу Pancrustacea. Положение Myriapoda, Chelicerata и Pancrustacea по состоянию на апрель 2012 г. остается неясным. В некоторых исследованиях Myriapoda сгруппированы с Chelicerata (образуя Myriochelata ); в других исследованиях Myriapoda сгруппирована с Pancrustacea (образуя Mandibulata ), или Myriapoda может быть сестрой Chelicerata plus Pancrustacea.
| традиционный Crustacea |
Размещение вымерших трилобитов также часто является предметом споров. Одна из новейших гипотез состоит в том, что хелицеры произошли от той же пары придатков, которые превратились в усики у предков Mandibulata, что поместило трилобитов с усиками ближе к Mandibulata, чем Chelicerata.
Поскольку Международный кодекс зоологической номенклатуры не признает никакого приоритета над рангом семьи, многие группы более высокого уровня могут упоминаться под разными именами.
 Насекомые и скорпионы, продаваемые в продуктовом ларьке в Бангкоке, Таиланде
Насекомые и скорпионы, продаваемые в продуктовом ларьке в Бангкоке, Таиланде Ракообразные, такие как крабы, омары, раки, креветки и креветки давно стали частью человеческой кухни и теперь выращиваются в коммерческих целях. Насекомые и их личинки по крайней мере так же питательны, как мясо, и их едят как в сыром, так и в приготовленном виде во многих культурах, но не в большинстве европейских, индуистских и исламских культур. Приготовленные тарантулы считаются деликатесом в Камбодже, а у индейцев пиароа южной Венесуэлы после очень раздражающих волос - паучьих волос. основная система защиты - снимаются. Люди также непреднамеренно съедают членистоногих в других пищевых продуктах, а правила безопасности пищевых продуктов устанавливают приемлемые уровни загрязнения для различных видов пищевых продуктов. Намеренное выращивание членистоногих и других мелких животных для употребления в пищу, именуемое мини-животноводством, теперь становится экологически обоснованной концепцией животноводства. Коммерческое разведение бабочек предоставляет стада чешуекрылых оранжереям, образовательным выставкам, школам, исследовательским центрам и культурным мероприятиям.
Однако наибольший вклад членистоногих в обеспечение человека продовольствием вносит опыление : в исследовании 2008 года было изучено 100 сельскохозяйственных культур, которые ФАО перечисляет как выращиваемые для производства продуктов питания, и оценивается экономическая стоимость опыления в 153 евро. миллиардов, или 9,5% стоимости мирового сельскохозяйственного производства, используемого для производства продуктов питания человека в 2005 году. Помимо опыления, пчелы производят мед, который является основой быстрорастущей отрасли и международного
Красный краситель. кошениль, полученная из центральноамериканского вида насекомых, была экономически важна для ацтеков и майя. В то время как регион находился под контролем Испании, он стал вторым по прибыльности экспортным товаром для Мексики и теперь восстанавливает некоторые позиции, которые он уступил синтетическим конкурентам. Shellac, смола, выделяемая видом насекомых, произрастающих в южной Азии, исторически использовалась в больших количествах для многих применений, в которых ее в основном заменяли синтетическими смолами, но она все еще используется в деревообработке и в качестве пищевой добавки. Кровь подковообразных крабов содержит свертывающий агент Limulus Amebocyte Lysate, который в настоящее время используется для проверки того, что антибиотики и почечные механизмы не содержат опасных бактерий, а также для выявления спинномозговой менингит и некоторые раковые заболевания. Судебная энтомология использует данные, полученные от членистоногих, для установления времени, а иногда и места смерти человека, а в некоторых случаях и причины. В последнее время насекомые также привлекли внимание как потенциальные источники лекарств и других лекарственных веществ.
Относительная простота строения тела членистоногих, позволяющая им перемещаться по различным поверхностям как на суше, так и в воде, сделала их можно использовать в качестве моделей для робототехники. Избыточность, обеспечиваемая сегментами, позволяет членистоногим и биомиметическим роботам нормально передвигаться даже с поврежденными или потерянными придатками.
| Заболевания | Насекомые | Случаев в год | Летальных исходов в год |
|---|---|---|---|
| Малярия | Anopheles комар | 267 M | 1-2 M |
| Лихорадка денге | Aedes комар | ? | ? |
| Желтая лихорадка | Aedes москит | 4,432 | 1,177 |
| Филяриоз | Culex комар | 250 M | неизвестно |
Хотя членистоногие - самый многочисленный тип на Земле, а тысячи видов членистоногих ядовиты, они наносят относительно мало серьезных укусов и укусов людей. Гораздо серьезнее воздействие на людей таких болезней, как малярия, переносимых кровососущими насекомыми. Другие кровососущие насекомые заражают домашний скот болезнями, которые убивают многих животных и значительно снижают полезность других. Клещи могут вызывать клещевой паралич и несколько паразитарных болезней в людях. Некоторые из близкородственных клещей также заражают людей, вызывая сильный зуд, а другие вызывают аллергические заболевания, включая сенную лихорадку, астму и экзема.
Многие виды членистоногих, в основном насекомые, но также и клещи, являются сельскохозяйственными и лесными вредителями. Клещ деструктор Варроа стал самой большой проблемой, с которой столкнулись пчеловоды во всем мире. Усилия по борьбе с членистоногими вредителями с помощью широкомасштабногоиспользования пестицидов оказали долгосрочное воздействие на здоровье человека и биоразнообразие. Повышение устойчивости членистоногих к пестицидам привело к разработке интегрированной борьбы с вредителями с использованием широкого спектра мер, включая биологический контроль. Хищные клещи могут быть полезны при борьбе с некоторыми клещами-вредителями.