
Войти
Биосинтез - многоступенчатый, ферментный - катализируемый процесс, где субстраты преобразуются в более сложные продукты в живых организмах. В биосинтезе простые соединения модифицируются, превращаются в другие соединения или соединяются вместе с образованием макромолекул. Этот процесс часто состоит из метаболических путей. Некоторые из этих биосинтетических путей расположены внутри одной клеточной органеллы, в то время как другие включают ферменты, которые расположены во множестве клеточных органелл. Примеры этих биосинтетических путей включают продукцию липидных мембран компонентов и нуклеотидов. Биосинтез обычно синонимичен с анаболизмом.
Необходимыми элементами для биосинтеза являются: соединения-предшественники, химическая энергия (например, ATP ), и каталитические ферменты, которым могут потребоваться коферменты (например, NADH, NADPH ). Эти элементы создают мономеры, строительные блоки для макромолекул. Некоторые важные биологические макромолекулы включают: белки, которые состоят из мономеров аминокислот, соединенных посредством пептидных связей, и молекул ДНК, которые являются состоит из нуклеотидов, соединенных фосфодиэфирными связями.
Биосинтез происходит из-засерия химических реакций. Для протекания этих реакций необходимы следующие элементы:
В простейшем смысле реакции, происходящие при биосинтезе, имеют следующий формат :
![{\displaystyle {\ce {Reactant ->[Фермент] Продукт}}}]( https://wikimedia.org/api/rest_v1/media/math/render/svg/d97fbef58f64fb0b7d05d90fa0a73b05f5919fc8 )
Ниже приведены некоторые варианты этого основного уравнения, которые будут обсуждаться более подробно ниже:

![{\displaystyle {\ce {{Precursor~molecule}+Cofactor->[фермент] макромолекула}}}]( https://wikimedia.org/api/rest_v1/media/math/render/svg/b192f25eec9ceebed179bfeb43ae27795f3a91be )

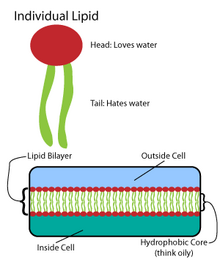 Липидный бислой мембраны
Липидный бислой мембраны Многие интриги Катовые макромолекулы синтезируются в виде простых повторяющихся структур. Например, простейшие структуры липидов - это жирные кислоты. Жирные кислоты представляют собой углеводородные производные; они содержат карбоксильную группу «голова» и углеводородную цепь «хвост». Эти жирные кислоты создают более крупные компоненты, которые, в свою очередь, включают нековалентные взаимодействия с образованием липидного бислоя. Цепи жирных кислот присутствуют в двух основных компонентах мембранных липидов: фосфолипидах и сфинголипидах. Третий основной компонент мембраны, холестерин, не содержит этих звеньев жирных кислот.
Основа всех биомембран состоит из бислоя структура фосфолипидов. Молекула фосфолипида амфипатическая ; он содержит гидрофильную полярную головку и гидрофобный неполярный хвост. Фосфолипидные головки взаимодействуют друг с другом и водной средой, тогда как углеводородные хвосты ориентируются в центре, вдали от воды. Эти последние взаимодействия управляют двухслойной структурой, которая действует как барьер для ионов и молекул.
Существуют различные типы фосфолипидов; следовательно, пути их синтеза различаются. Однако первая стадия синтеза фосфолипидов включает образование фосфатидата или диацилглицерин-3-фосфата в эндоплазматическом ретикулуме и внешней мембране митохондрий. Путь синтеза приведен ниже:

Путь начинается с глицерин-3-фосфата, который превращается в лизофосфатидат посредством добавления цепи жирной кислоты, обеспечиваемой ацилкоферментом A. Затем лизофосфатидат превращается в фосфатидат посредством добавления другой цепи жирной кислоты, вносимой вторым ацил-КоА; все эти стадии катализируются ферментом глицеринфосфат ацилтрансфераза. Синтез фосфолипидов продолжается в эндоплазматическом ретикулуме, и пути биосинтеза расходятся в зависимости от компонентов конкретного фосфолипида.
Как и фосфолипиды, эти производные жирных кислот имеют полярную головку и неполярные хвосты. В отличие от фосфолипидов, сфинголипиды имеют основу сфингозина. Сфинголипиды существуют в эукариотических клетках, и их особенно много в центральной нервной системе. Например, сфингомиелин является частью миелиновой оболочки нервных волокон.
Сфинголипиды образуются из церамидов, которые состоят из цепи жирной кислоты, присоединенной к аминогруппе сфингозиновый остов. Эти церамиды синтезируются путем ацилирования сфингозина. Путь биосинтеза сфингозина представлен ниже:

Как показано на изображении, во время синтеза сфингозина пальмитоил-КоА и серин подвергаются реакции конденсации, которая приводит к образованию дегидросфингозина. Затем этот продукт восстанавливается с образованием дигидроспингозина, который превращается в сфингозин посредством реакции окисления посредством FAD.
Этот липид принадлежит к группе класс молекул, называемых стеринами. Стерины имеют четыре конденсированных кольца и гидроксильную группу. Холестерин - особенно важная молекула. Он не только является компонентом липидных мембран, но также является предшественником нескольких стероидных гормонов, включая кортизол, тестостерон и эстроген..
Холестерин синтезируется из ацетил-КоА. Путь показан ниже:

В более общем смысле этот синтез происходит в три стадии, первая стадия происходит в цитоплазме, а вторая и третья стадии - в эндоплазматическом ретикулуме. Этапы следующие:
Биосинтез нуклеотидов включает реакции, катализируемые ферментами, которые превращают субстраты в более сложные продукты. Нуклеотиды являются строительными блоками ДНК и РНК. Нуклеотиды состоят из пятичленного кольца, образованного из рибозы сахара в РНК и дезоксирибозного сахара в ДНК; эти сахара связаны с пуриновым или пиримидиновым основанием с гликозидной связью и фосфатной группой в положении 5 ' сахара.
 Синтез IMP .
Синтез IMP .Нуклеотиды ДНК аденозин и гуанозин состоят из пуринового основания, присоединенного к сахар рибозы с гликозидной связью. В случае нуклеотидов РНК дезоксиаденозин и дезоксигуанозин пуриновые основания присоединены к сахару дезоксирибозы гликозидной связью. Пуриновые основания на нуклеотидах ДНК и РНК синтезируются по двенадцатиступенчатому механизму реакции, присутствующему в большинстве одноклеточных организмов. Высшие эукариоты используют аналогичный механизм реакции из десяти стадий реакции. Пуриновые основания синтезируют путем превращения фосфорибозилпирофосфата (PRPP) в инозинмонофосфат (IMP), который является первым ключевым промежуточным соединением в биосинтезе пуриновых оснований. Дальнейшая ферментативная модификация IMP дает аденозиновые и гуанозиновые основания нуклеотидов.
 Уридинмонофосфат (UMP) биосинтез
Уридинмонофосфат (UMP) биосинтез Другие нуклеотидные основания ДНК и РНК, которые связаны с рибозным сахаром через гликозидную связь: тимин, цитозин и урацил (который встречается только в РНК). В биосинтезе уридинмонофосфата участвует фермент, расположенный на внутренней мембране митохондрий, и многофункциональные ферменты, расположенные в цитозоле.
После синтеза уридинового нуклеотидного основания, другие основания, цитозин и тимин синтезируются. Биосинтез цитозина - это двухэтапная реакция, которая включает превращение UMP в UTP. Добавление фосфата к UMP катализируется ферментом киназой. Фермент CTP-синтаза катализирует следующую стадию реакции: превращение UTP в CTP путем переноса аминогруппы с глутамина на уридин; это формирует цитозиновую основу CTP. Механизм, который описывает реакцию UTP + ATP + глутамин ⇔ CTP + ADP + глутамат, приведен ниже:

Цитозин - это нуклеотид, который присутствует как в ДНК, так и в РНК. Однако урацил находится только в РНК. Следовательно, после синтеза УТФ он должен быть преобразован в дезокси форму для включения в ДНК. В этом превращении участвует фермент рибонуклеозидтрифосфатредуктаза. На эту реакцию, которая удаляет 2'-ОН сахара рибозы с образованием дезоксирибозы, не влияют основания, присоединенные к сахару. Эта неспецифичность позволяет рибонуклеозидтрифосфатредуктазе преобразовывать все нуклеотидтрифосфаты в дезоксирибонуклеотид по аналогичному механизму.
В отличие от урацила, тиминовые основания обнаруживаются в основном в ДНК., а не РНК. Клетки обычно не содержат тиминовых оснований, которые связаны с сахарами рибозы в РНК, что указывает на то, что клетки синтезируют только связанный с дезоксирибозой тимин. Фермент тимидилатсинтетаза отвечает за синтез остатков тимина от dUMP до dTMP. Эта реакция переносит метил группу на урациловое основание dUMP с образованием dTMP. Реакция тимидилатсинтазы, dUMP + 5,10-метилентетрагидрофолат ⇔ dTMP + дигидрофолат, показана справа.
Хотя существуют различия между эукариотами и синтез прокариотической ДНК, в следующем разделе обозначены ключевые характеристики репликации ДНК, общие для обоих организмов.
ДНК состоит из нуклеотидов, которые соединены фосфодиэфирными связями. Синтез ДНК, который происходит в ядре, является полуконсервативным процессом, что означает, что полученная молекула ДНК содержит исходную цепь из родительской структуры и новую цепь. Синтез ДНК катализируется семейством ДНК-полимераз, для которых требуются четыре дезоксинуклеозидтрифосфата, матричная цепь и праймер со свободным 3'OH, в котором включают нуклеотиды.
Для репликации ДНК репликационная вилка создается ферментами, называемыми геликазами, которые раскручивают спираль ДНК. Топоизомеразы на репликационной вилке удалите суперспирали, вызванные раскручиванием ДНК, и белки, связывающие одноцепочечную ДНК, поддерживают две матрицы одноцепочечной ДНК, стабилизированные перед репликацией.
Синтез ДНК инициируется РНК-полимеразой примазой, которая создает праймер РНК со свободным 3'OH. Этот праймер прикреплен к одноцепочечной ДНК-матрице, и ДНК-полимераза удлиняет цепь за счет включения нуклеотидов; ДНК-полимераза также проверяет вновь синтезированную цепь ДНК.
Во время реакции полимеризации, катализируемой ДНК-полимеразой, нуклеофильная атака происходит со стороны 3'OH растущей цепи на самый внутренний атом фосфора дезоксинуклеозидтрифосфат; это приводит к образованию фосфодиэфирного мостика, который присоединяет новый нуклеотид и высвобождает пирофосфат.
Два типа цепей создаются одновременно во время репликации: ведущая цепь, которая является синтезируется непрерывно и растет по направлению к вилке репликации, а также к отстающей цепи, которая прерывисто образуется во фрагментах Окадзаки и отрастает от вилки репликации. Фрагменты Окадзаки ковалентно соединены ДНК-лигазой с образованием непрерывной цепи. Затем для завершения репликации ДНК удаляются праймеры РНК, а образовавшиеся промежутки заменяются ДНК и соединяются с помощью ДНК-лигазы.
Белок представляет собой полимер, состоящий из амино кислоты, связанные пептидными связями. В природе обнаружено более 300 аминокислот, из которых только двадцать, известные как стандартные аминокислоты, являются строительными блоками для белка. Только зеленые растения и большинство микробов способны синтезировать все 20 стандартных аминокислот, которые необходимы всем живым видам. Млекопитающие могут синтезировать только десять из двадцати стандартных аминокислот. Другие аминокислоты, валин, метионин, лейцин, изолейцин, фенилаланин, лизин, треонин и триптофан для взрослых и гистидин и аргинин для младенцев представляют с пищей.
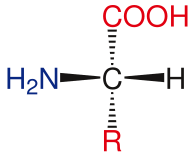 L-аминокислота
L-аминокислота Общая структура аминокислот включает первичную аминогруппу, карбоксильную группу и функциональную группу присоединен к α-ограничоду. Различные аминокислоты идентифицируются по функциональной группе. В результате трех различных групп, присоединенных к α-уровню, аминокислоты представляют собой асимметричные молекулы. Для всех стандартных аминокислот, кроме глицина, -углерод является хиральным центром. В случае глицина α-углеродный два атома водорода, что симметрии этой молекуле. За исключением пролина, все аминокислоты, обнаруженные в жизни, имеют конформацию L-изоформы. Пролин имеет функциональную группу на α-кольцо, который образует с аминогруппой.

Один из основных этапов биосинтеза аминокислоты включает включение группы азота на α-углерод. В клетках есть два основных пути включения азота. Один путь включает фермент глутаминоксоглутарат аминотрансферазу (GOGAT), который удаляет амидную группу из глутамина и переносит ее на 2-оксоглутарат, производящие две молекулы глутамата. В этой реакции катализа глутамин служит средством азота. Изображение, изображающее эту реакцию, находится справа.
Другой путь включения азота в α-углерод включает фермент глутаматдегидрогеназу (GDH). GDH способен переносить аммиак на 2-оксоглутарат и образовывать глутамат. Кроме фермент глутаминсинтетаза (GS) способен переносить аммиак на глутамат и синтезировать глутамин, пополняя запасы глутамина.
Семейство глутаминовых аминокислот включает аминокислоты, производные от аминокислоты глутамата. Это семейство включает: глутамат, глутамин, пролин и аргинин. Это семейство также включает аминокислоты лизин, которая является производным α-кетоглутарата.
. Биосинтез глутамата и глутамина является ключевым этапом в усвоении азота, описанном выше. Ферменты GOGAT и GDH катализируют реакции ассимиляции азота.
В бактериях фермент глутамат-5-киназа запускает биосинтез пролина путем переноса фосфатной группы с АТФ на глутамат. Следующая реакция катализируется ферментом пирролин-5-карбоксилатсинтазой (P5CS), который катализирует восстановление ϒ-карбоксильной группы L-глутамат-5-фосфата. Это приводит к образованию полуальдегида глутамата, который спонтанно циклически превращается в пирролин-5-карбоксилат. Пирролин-5-карбоксил дополнительно восстанавливается ферментом пирролин-5-карбоксилатредуктазой (P5CR) с образованием аминокислоты пролина.
На первой стадии биосинтеза аргинина у бактерий глутамат ацетилируется переносом ацетильной группы из ацетил-КоА в положение N-α; это предотвращает самопроизвольную циклизацию. Фермент N-ацетилглутаматсинтаза (глутамат-N-ацетилтрансфераза) отвечает за катализ на стадии ацетилирования. Последующие стадии катализируются ферментами N-ацетилглутаматкиназа, N-ацетил-гамма-глутамилфосфатредуктаза и ацетилорнитин / сукцинилдиаминопимелатаминотрансфераза и N-ацетил-L-орнитин. Ацетильная группа ацетилорнитина удаляется ферментом ацетилорнитиназой (AO) или орнитинацетилтрансферазой (OAT), и это дает орнитин. Затем ферменты цитруллин и аргининосукцинат превращают орнитин в аргинин.
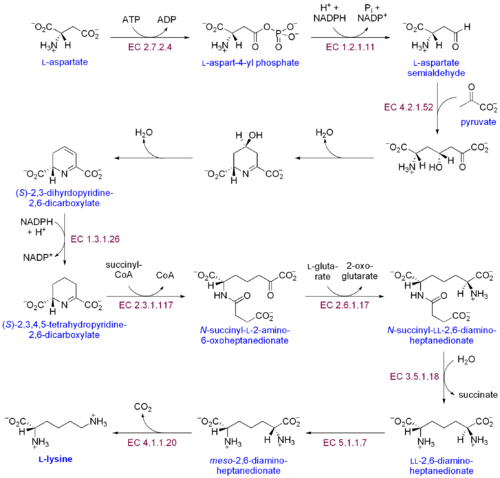 Путь диаминопимелиновой кислоты
Путь диаминопимелиновой кислоты Существует два различных пути биосинтеза лизина: путь диаминопимелиновой кислоты и α-аминоадипатный путь. Наиболее распространенным из двух путей синтеза является путь диаминопимелиновой кислоты; Он инициирует путь диаминопимелиновой кислоты посредством фосфорилирования аспартата и образования аспартилфосфата.
Семейство сериновых аминокислот включает: серин, цистеин и глицин. Большинство микроорганизмов и методов получают серу для синтеза метионина из аминокислот цистеина. Кроме того, которое превращает серина в глицин кристалл, необходимые для биосинтеза метионина и гистидина.
Во время биосинтеза серина фермент фосфоглицератдегидрогеназа катализирует начальную реакцию, окисляет 3 -фосфо-D-глицерат с получением 3-фосфонооксипирувата. Следующая катализируется ферментом фосфосерин-аминотрансферазой, который переносит аминогруппу с глутамата на 3-фосфоноксипируват с реакцией образования L-фосфосерина. Последний этап катализируется ферментом фосфосеринфосфатазой, который дефосфорилирует L-фосфосерин с образованием L-серина.
. Существует два известных пути биосинтеза глицина. Организмы, которые используют этанол и ацетат в качестве основного источника углерода, используют гликонеогенный путь для синтеза глицина. Другой путь биосинтеза глицина как известен гликолитический путь. Этот путь превращает серин, синтезированный из промежуточных продуктов гликолиза, в глицин. В гликолитическом пути фермент серингидроксиметилтрансфераза катализирует расщепление серина с глицина и переносит расщепленную углеродную группу серина на тетрагидрофолат с образованием 5,10-метилен-тетрагидрофолат.
Биосинтез цистеина представляет собой двухстадийную реакцию, которая включает включение неорганической серы. У микроорганизмов и растений фермент серинацетилтрансфераза катализирует перенос ацетильной группы от ацетил-КоА на L-серин с образованием O-ацетил-L-серина. На следующей стадии реакции, катализируемым ферментом O-ацетилсерин (тиол) лиазой, ацетильная группа O-ацетил-L-серина заменяется сульфидом с образованием цистеина.
Семейство аспартата аминокислоты включает: треонин, лизин, метионин, изолейцин и аспартат. Лизин и изолейцин считаются частью семьи аспартатов, хотя часть их углеродного скелета происходит от пирувата. В случае метионина металлический организм является производным серина и серной группы, но организмов он является производным цистеина.
Биосинтез аспартата представляет собой одностадийную реакцию, которая катализируется одним фермент. Фермент аспартатамирансфераза катализирует перенос аминогруппы от аспартата на α-кетоглутарат с образованием глутамата и оксалоацетата. Аспарагин синтезируется АТФ-зависимым присоединением аминогруппы к аспартату; аспарагинсинтетаза катализирует добавление азота из глутамина или растворимого аммиака к аспартату с образованием аспарагина.
 Путь биосинтеза лизина диаминопимелиновой кислоты
Путь биосинтеза лизина диаминопимелиновой кислоты Путь биосинтеза лизина диаминопимелиновой кислоты принадлежит к семейству аспартатов аминогруппы. кислоты. Этот путь включает девять катализируемых ферментами факторов, превращающие их в лизин.
 Антикодон тРНК взаимодействует с кодоном мРНК, чтобы связывают аминокислоты с растущей полипептидной цепью.
Антикодон тРНК взаимодействует с кодоном мРНК, чтобы связывают аминокислоты с растущей полипептидной цепью. 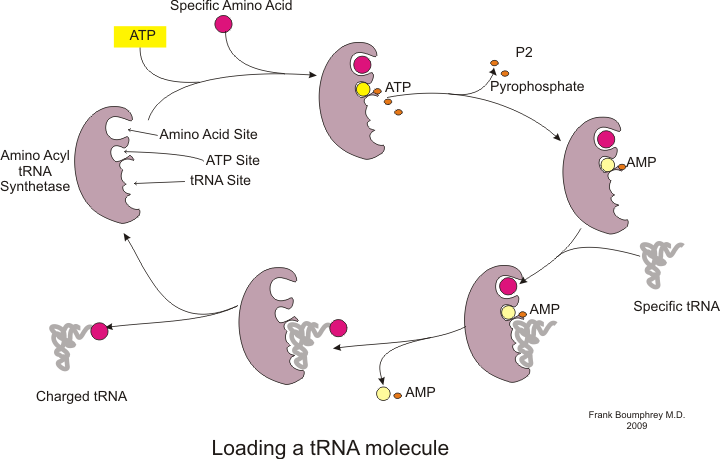 Процесс зарядки тРНК
Процесс зарядки тРНК Синтез белка происходит посредством процесса, называемого трансляцией. Во время трансляции генетический материал, называемый мРНК, считывается рибосомами с образованием цепи белка полипептида. Этот процесс требует транспортной РНК (тРНК), которая служит адаптером, связывая аминокислоты на одном конце и взаимодействуя с мРНК на другом конце; последнее спаривание тРНК и мРНК гарантирует, что к цепи добавлена правильная аминокислота. Синтез белка происходит в три фазы: инициация, удлинение и завершение. Прокариотическая трансляция отличается от эукариотической трансляции ; однако в этом разделе основное внимание будет уделено общему между двумя организмами.
Прежде чем можно будет начать трансляцию, должен произойти процесс связывания конкретной аминокислоты с соответствующей ей тРНК. Эта реакция, называемая зарядкой тРНК, катализируется аминоацил тРНК синтетазой. Определенная тРНК-синтетаза отвечает за распознавание и зарядку определенной аминокислоты. Кроме того, у этого фермента есть специальные дискриминаторные области, обеспечивающие правильное связывание тРНК с родственной ей аминокислотой. Первым шагом для присоединения аминокислоты к соответствующей тРНК является образование аминоацил-АМФ:

За этим следует перенос аминоацильной группы от аминоацил-AMP к молекуле тРНК. полученная молекула представляет собой аминоацил-тРНК :

Сочетание этих двух шагов, оба из которых катализируются аминоацил тРНК синтетазой, дает заряженную тРНК, готовую к добавлению аминокислот к растущей полипептидной цепи.
Помимо связывания аминокислоты, тРНК имеет трехнуклеотидную единицу, называемую антикодоном, которая со специфическими триплетами нуклеотидов на мРНК, называемыми кодонами ; кодоны кодируют конкретную аминокислоту. Это взаимодействие возможно благодаря рибосоме, которая служит местом синтеза белка. Рибосома имеет три сайта связывания тРНК: аминоацильный сайт (сайт A), пептидильный сайт (сайт P) и сайт выхода (сайт E).
В транскрипте мРНК содержится множество кодонов, и он очень часто аминокислота определяется более чем одним кодоном; это явление называется вырождением. Всего существует 64 кодона, по 61 каждого кода для одной из 20 аминокислот, в то время как оставшиеся кодоны определяют терминацию цепи.
Как упоминалось ранее, трансляция происходит в три фазы: начало, удлинение и завершение.
Завершение фазы инициации зависит от следующих трех событий:
1. Привлечение рибосомы к мРНК
2. Связывание заряженной тРНК инициатора с Р-сайтом рибосомы
3. Правильное выравнивание рибосомы со стартовым кодоном мРНК
После инициации полипептидная цепь удлиняется посредством взаимодействий антикодон: кодон, при этом рибосома добавляет аминокислоты к полипептидной цепи один за раз. Чтобы гарантировать правильное добавление аминокислот, необходимо выполнить следующие шаги:
1. Связывание правильной тРНК с сайтом A рибосомы
2. Образование пептидной связи между тРНК в сайте A и полипептидной цепью, присоединенной к тРНК в сайте P
3. Транслокация или продвижение комплекса тРНК-мРНК на три нуклеотида
Транслокация «запускает» тРНК на сайте E и сдвигает тРНК с сайта A на сайт P, оставляя Сайт, свободный для входящей тРНК, чтобы добавить другую аминокислоту.
Последний этап трансляции происходит, когда стоп-кодон попадает на сайт A. Затем выполняются следующие шаги:
1. Распознавание кодонов факторами высвобождения, что вызывает гидролиз полипептидной цепи из тРНК, расположенной в сайте P
2. Высвобождение полипептидной цепи
3. Диссоциация и «повторное использование» рибосомы для будущих процессов трансляции
Сводная таблица ключевых участников перевода приведена ниже:
| Ключевые участники перевода | Стадия перевода | Цель |
|---|---|---|
| тРНК-синтетаза | до инициации | Ответственная за заряд тРНК |
| мРНК | инициация, элонгация, терминация | Матрица для синтез белка; содержит области, названные кодонами, которые кодируют аминокислоты |
| тРНК | инициация, элонгация, терминация | Связывает сайты рибосом A, P, E; пары оснований антикодона с кодоном мРНК для включения правильных аминокислот в растущую полипептидную цепь |
| рибосома | инициация, удлинение, терминация | Направляет синтез белка и катализирует образование пептидной связи |
 Семейная гиперхолестеринемия вызывает отложения холестерина
Семейная гиперхолестеринемия вызывает отложения холестерина Ошибки в путях биосинтеза могут иметь пагубные последствия, включая неправильное формирование, макромолекул или недостаточное производство функциональных молекул. Ниже приведены примеры, показывающие сбои, демонстрирующие из-за такой неэффективности.