
Войти
Нейронные колебания или мозговые волны, представляют собой ритмичные или повторяющиеся паттерны нейронной активности в центральной нервной системе. Нервная ткань может генерировать осцилляторную активность многими способами, управляемыми либо механизмами внутри отдельных нейронов, либо взаимодействиями между нейронами. В отдельных нейронах колебания могут проявляться либо в виде колебаний мембранного потенциала, либо в виде ритмических паттернов потенциалов действия, которые затем вызывают колебательную активацию постсинаптических нейронов. На уровне нейронных ансамблей синхронизированная активность большого количества нейронов может вызывать макроскопические колебания, которые можно наблюдать на электроэнцефалограмме. Колебательная активность в группах нейронов обычно возникает в результате обратных связей между нейронами, которые приводят к синхронизации их паттернов возбуждения. Взаимодействие между нейронами может вызывать колебания с частотой, отличной от частоты срабатывания отдельных нейронов. Хорошо известным примером макроскопических нейронных колебаний является альфа-активность.
Нервные колебания наблюдались исследователями еще в 1924 году (Гансом Бергером ). Более чем через 50 лет внутреннее колебательное поведение было обнаружено в нейронах позвоночных, но его функциональная роль до сих пор полностью не изучена. Возможные роли нейронных колебаний включают в себя связывание признаков, механизмы передачи информации и формирование ритмического моторного выхода. За последние десятилетия было получено больше информации, особенно в области визуализации мозга. Основная область исследований в неврологии включает определение того, как генерируются колебания и каковы их роли. Колебательная активность в мозге широко наблюдается на разных уровнях организации и, как считается, играет ключевую роль в обработке нейронной информации. Многочисленные экспериментальные исследования подтверждают функциональную роль нервных колебаний; Однако единого толкования пока нет.
 Моделирование нейронных колебаний на частоте 10 Гц. Верхняя панель показывает пики отдельных нейронов (каждая точка представляет индивидуальный потенциал действия в популяции нейронов), а нижняя панель потенциал локального поля, отражающая их суммарную активность. На рисунке показано, как синхронизированные образцы потенциалов действия могут привести к макроскопическим колебаниям, которые можно измерить за пределами кожи головы.
Моделирование нейронных колебаний на частоте 10 Гц. Верхняя панель показывает пики отдельных нейронов (каждая точка представляет индивидуальный потенциал действия в популяции нейронов), а нижняя панель потенциал локального поля, отражающая их суммарную активность. На рисунке показано, как синхронизированные образцы потенциалов действия могут привести к макроскопическим колебаниям, которые можно измерить за пределами кожи головы. Ричард Кейтон обнаружил электрическую активность в мозговых полушариях кроликов и обезьян и представил свои открытия в 1875 году. Адольф Бек опубликовал в 1890 году свои наблюдения спонтанной электрической активности мозга кроликов и собак, которые включали ритмические колебания, измененные светом, обнаруживаемым непосредственно с помощью электродов. размещается на поверхности мозга. До Ганса Бергера Владимир Владимирович Правдич-Неминский опубликовал первую ЭЭГ животных и вызванный потенциал собаки.
Нейронные колебания наблюдаются по всей центральной нервной системе на всех уровнях, и включают последовательности импульсов, потенциалы локального поля и крупномасштабные колебания, которые можно измерить с помощью электроэнцефалографии (ЭЭГ). В общем, колебания могут быть охарактеризованы их частотой, амплитудой и фазой. Эти свойства сигнала могут быть извлечены из нейронных записей с помощью частотно-временного анализа. При крупномасштабных колебаниях считается, что изменения амплитуды являются результатом изменений синхронизации в пределах нейронного ансамбля , также называемой локальной синхронизацией. Помимо локальной синхронизации, может синхронизироваться колебательная активность отдаленных нервных структур (одиночных нейронов или нейронных ансамблей). Нейронные колебания и синхронизация были связаны со многими когнитивными функциями, такими как передача информации, восприятие, моторный контроль и память.
Нейронные колебания наиболее широко изучались в нейронной активности, генерируемой большими группами нейронов. Масштабную активность можно измерить с помощью таких методов, как ЭЭГ. В целом, сигналы ЭЭГ имеют широкий спектр спектра, аналогичный розовому шуму, но также обнаруживают колебательную активность в определенных частотных диапазонах. Первым обнаруженным и наиболее известным частотным диапазоном является альфа-активность (8–12 Гц ), которую можно обнаружить в затылочной доле во время расслабленного бодрствования и которая усиливается. когда глаза закрыты. Другие диапазоны частот: дельта (1–4 Гц), тета (4–8 Гц), бета (13–30 Гц), диапазоны частот с низкой гаммой (30–70 Гц) и высокой гаммой (70–150 Гц), где более быстрые ритмы, такие как гамма-активность, связаны с когнитивной обработкой. Действительно, сигналы ЭЭГ резко меняются во время сна и показывают переход от более быстрых частот к более медленным частотам, таким как альфа-волны. Фактически, разные стадии сна обычно характеризуются спектральным содержанием. Следовательно, нейронные колебания были связаны с когнитивными состояниями, такими как осознание и сознание.
. Хотя нейронные колебания активности человеческого мозга в основном исследуются с помощью записей ЭЭГ, они также наблюдаются с помощью более инвазивной записи. такие методы, как единичные записи. Нейроны могут генерировать ритмические паттерны потенциалов действия или пиков. Некоторые типы нейронов имеют тенденцию срабатывать на определенных частотах, так называемые резонаторы. Взрыв - еще одна форма ритмических пиков. Шаблоны пиков считаются фундаментальными для кодирования информации в мозгу. Колебательная активность также может наблюдаться в виде колебаний подпорогового мембранного потенциала (т.е. в отсутствие потенциалов действия). Если многочисленные нейроны вспыхивают в синхронности, они могут вызвать колебания потенциалов локального поля. Количественные модели позволяют оценить силу нейронных колебаний в записанных данных.
Нейронные колебания обычно изучаются с математической основы и относятся к области «нейродинамики», области исследований в когнитивных науках, в котором большое внимание уделяется динамическому характеру нейронной активности при описании функции мозга. Он рассматривает мозг как динамическую систему и использует дифференциальные уравнения для описания эволюции нейронной активности во времени. В частности, он направлен на то, чтобы связать динамические модели активности мозга с когнитивными функциями, такими как восприятие и память. В очень абстрактной форме нейронные колебания можно анализировать аналитически. При изучении в более физиологически реалистичных условиях осцилляторная активность обычно изучается с помощью компьютерного моделирования из вычислительной модели.
Функции нейронных колебаний имеют широкий диапазон и различаются для разных типов колебательной активности.. Примерами являются создание ритмической активности, такой как сердцебиение и нейронная привязка сенсорных функций при восприятии, таких как форма и цвет объекта. Нервные колебания также играют важную роль во многих неврологических расстройствах, таких как чрезмерная синхронизация во время приступа активности при эпилепсии или тремора у пациентов с Болезнь Паркинсона. Колебательная активность также может использоваться для управления внешними устройствами в интерфейсах мозг-компьютер, в которых субъекты могут управлять внешним устройством, изменяя амплитуду определенной ритмики мозга.
Колебательная активность наблюдается во всей центральной нервной системе на всех уровнях организации. Широко известны три различных уровня: микромасштаб (активность отдельного нейрона), мезомасштаб (активность локальной группы нейронов) и макромасштаб (активность разных областей мозга).
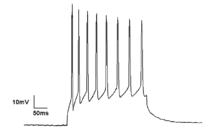 Тоник образец возбуждения одиночного нейрона, демонстрирующего ритмическую импульсную активность
Тоник образец возбуждения одиночного нейрона, демонстрирующего ритмическую импульсную активность Нейроны генерируют потенциалы действия в результате изменений электрического мембранного потенциала. Нейроны могут последовательно генерировать несколько потенциалов действия, образуя так называемые цепочки шипов. Эти последовательности импульсов являются основой нейронного кодирования и передачи информации в мозгу. Цепочки шипов могут образовывать всевозможные паттерны, такие как ритмические всплески и разрывные, и часто демонстрируют колебательную активность. Колебательную активность в отдельных нейронах также можно наблюдать в подпороговых колебаниях мембранного потенциала. Эти ритмические изменения мембранного потенциала не достигают критического порога и, следовательно, не приводят к возникновению потенциала действия. Они могут быть результатом постсинаптических потенциалов от синхронных входов или внутренних свойств нейронов.
Нейронные всплески можно классифицировать по моделям их активности. Возбудимость нейронов можно подразделить на классы I и II. Нейроны класса I могут генерировать потенциалы действия с произвольно низкой частотой в зависимости от входной мощности, тогда как нейроны класса II генерируют потенциалы действия в определенной полосе частот, которая относительно нечувствительна к изменениям входной силы. Нейроны класса II также более склонны к подпороговым колебаниям мембранного потенциала.
Группа нейронов также может генерировать колебательную активность. Посредством синаптических взаимодействий паттерны возбуждения разных нейронов могут стать синхронизированными, и ритмические изменения электрического потенциала, вызванные их потенциалами действия, будут складываться (конструктивная интерференция ). То есть синхронизированные схемы возбуждения приводят к синхронизированному входу в другие области коры, что вызывает колебания большой амплитуды потенциала локального поля. Эти крупномасштабные колебания также можно измерить за пределами кожи головы с помощью электроэнцефалографии (ЭЭГ) и магнитоэнцефалографии (МЭГ). Электрические потенциалы, генерируемые отдельными нейронами, слишком малы, чтобы их можно было уловить за пределами кожи головы, а активность ЭЭГ или МЭГ всегда отражает совокупность синхронной активности тысяч или миллионов нейронов, имеющих аналогичную пространственную ориентацию. Нейроны в нейронном ансамбле редко все активируются в один и тот же момент, то есть полностью синхронизированы. Вместо этого вероятность срабатывания ритмически модулируется таким образом, что нейроны с большей вероятностью срабатывают одновременно, что вызывает колебания их средней активности (см. Рисунок вверху страницы). Таким образом, частота крупномасштабных колебаний не обязательно должна соответствовать шаблону возбуждения отдельных нейронов. Изолированные нейроны коры головного мозга регулярно срабатывают при определенных условиях, но в неповрежденном мозге корковые клетки подвергаются бомбардировке сильно колеблющимся синаптическим входом и, как правило, срабатывают случайно. Однако, если вероятность появления большой группы нейронов ритмически модулируется с общей частотой, они будут генерировать колебания в среднем поле (см. Также рисунок вверху страницы). Нейронные ансамбли могут генерировать колебательную активность эндогенно посредством локальных взаимодействий между возбуждающими и тормозящими нейронами. В частности, тормозящие интернейроны играют важную роль в обеспечении синхронности нейронного ансамбля, создавая узкое окно для эффективного возбуждения и ритмически модулируя скорость возбуждения возбуждающих нейронов.
Нейронные колебания также могут возникать в результате взаимодействия между различными областями мозга, связанными через структурный коннектом. Временные задержки играют здесь важную роль. Поскольку все области мозга двунаправленно связаны, эти связи между областями мозга образуют петли обратной связи. Положительная обратная связь, как правило, вызывает колебательную активность, частота которой обратно пропорциональна времени задержки. Примером такой петли обратной связи являются связи между таламусом и корой - таламокортикальными излучениями. Эта таламокортикальная сеть способна генерировать колебательную активность, известную как повторяющийся таламо-кортикальный резонанс. Таламокортикальная сеть играет важную роль в генерации альфа-активности. В модели сети всего мозга с реалистичной анатомической связностью и задержками распространения между областями мозга колебания в бета-диапазоне возникают из-за частичной синхронизации подмножеств областей мозга, колеблющихся в гамма-диапазоне (генерируемых в мезоскопический уровень).
Ученые определили некоторые внутренние свойства нейронов, которые играют важную роль в генерации колебаний мембранного потенциала. В частности, потенциалзависимые ионные каналы имеют решающее значение для генерации потенциалов действия. Динамика этих ионных каналов отражена в хорошо известной модели Ходжкина-Хаксли, которая описывает, как потенциалы действия инициируются и распространяются с помощью набора дифференциальных уравнений. Используя бифуркационный анализ, можно определить различные колебательные разновидности этих нейрональных моделей, что позволяет классифицировать типы нейронных ответов. Осцилляторная динамика спайков нейронов, идентифицированная в модели Hodgkin-Huxley, полностью согласуется с эмпирическими данными. Помимо периодических всплесков, подпороговые колебания мембранного потенциала, то есть резонансное поведение, которое не приводит к появлению потенциалов действия, также могут способствовать колебательной активности, способствуя синхронной активности соседних нейронов. Подобно нейронам-пейсмекерам в генераторах центральных паттернов, подтипы кортикальных клеток запускают всплески спайков (короткие группы спайков) ритмично с предпочтительными частотами. Разрывные нейроны могут служить в качестве кардиостимуляторов для синхронных сетевых колебаний, а всплески импульсов могут лежать в основе или усиливать нейронный резонанс.
Помимо внутренних свойств нейронов, биологические Свойства нейронной сети также являются важным источником колебательной активности. Нейроны связываются друг с другом через синапсы и влияют на синхронизацию последовательности спайков в постсинаптических нейронах. В зависимости от свойств соединения, таких как сила связи, временная задержка и от того, является ли соединение возбуждающим или тормозным, последовательности импульсов взаимодействующих нейронов могут стать синхронизированными. Нейроны локально связаны, образуя небольшие кластеры, которые называются нейронными ансамблями. Определенные сетевые структуры способствуют колебательной активности на определенных частотах. Например, нейронная активность, генерируемая двумя популяциями взаимосвязанных тормозных и возбуждающих клеток, может демонстрировать спонтанные колебания, которые описываются моделью Уилсона-Коуэна.
. Если группа нейронов участвует в синхронизированной колебательной активности, нейронный ансамбль может быть математически представлен как единственный осциллятор. Различные нейронные ансамбли связаны между собой дальнодействующими связями и образуют сеть слабосвязанных осцилляторов следующего пространственного масштаба. Слабосвязанные осцилляторы могут генерировать ряд динамических сигналов, включая колебательную активность. Связи на больших расстояниях между различными структурами мозга, такими как таламус и кора (см. таламокортикальные колебания ), связаны с задержками по времени из-за конечного скорость проводимости аксонов. Поскольку большинство соединений являются взаимными, они образуют петли обратной связи, которые поддерживают колебательную активность. Колебания, зарегистрированные в нескольких областях коры головного мозга, могут быть синхронизированы с образованием крупномасштабных мозговых сетей, динамику и функциональную взаимосвязь которых можно изучить с помощью спектрального анализа и причинности Грейнджера меры. Согласованная активность крупномасштабной мозговой активности может формировать динамические связи между областями мозга, необходимыми для интеграции распределенной информации.
В дополнение к быстрым прямым синаптическим взаимодействиям между В нейронах, образующих сеть, осцилляторная активность регулируется нейромодуляторами в гораздо более медленном масштабе времени. То есть известно, что уровни концентрации определенных нейротрансмиттеров регулируют уровень колебательной активности. Например, было показано, что концентрация ГАМК положительно коррелирует с частотой колебаний индуцированных стимулов. Ряд ядер в стволе мозга имеют диффузные выступы по всему мозгу, влияющие на уровни концентрации нейромедиаторов, таких как норэпинефрин, ацетилхолин и серотонин. Эти системы нейротрансмиттеров влияют на физиологическое состояние, например, бодрствование или возбуждение, и оказывают выраженное влияние на амплитуду различных мозговых волн, таких как альфа-активность.
Колебания часто можно описать и проанализировать с помощью математики. Математики выделили несколько динамических механизмов, порождающих ритмичность. Среди наиболее важных - гармонические (линейные) генераторы, генераторы предельного цикла и генераторы с задержкой обратной связи. Гармонические колебания возникают в природе очень часто - например, звуковые волны, движение маятника и всевозможные колебания. Обычно они возникают, когда физическая система в небольшой степени возмущена из состояния с минимальной энергией, и хорошо понимаются математически. Управляемые шумом гармонические осцилляторы реалистично имитируют альфа-ритм в ЭЭГ бодрствования, а также медленные волны и веретена в ЭЭГ во сне. Успешные алгоритмы анализа ЭЭГ были основаны на таких моделях. Некоторые другие компоненты ЭЭГ лучше описываются колебаниями предельного цикла или задержанной обратной связи. Колебания предельного цикла возникают из-за физических систем, которые показывают большие отклонения от равновесия, тогда как колебания с запаздыванием возникают, когда компоненты системы влияют друг на друга после значительных временных задержек. Колебания предельного цикла могут быть сложными, но есть мощные математические инструменты для их анализа; математика колебаний с запаздыванием по сравнению с этим примитивна. Линейные осцилляторы и осцилляторы ограниченного цикла качественно различаются по тому, как они реагируют на колебания входного сигнала. В линейном генераторе частота более или менее постоянна, но амплитуда может сильно варьироваться. В генераторе с предельным циклом амплитуда имеет тенденцию быть более или менее постоянной, но частота может сильно варьироваться. сердцебиение является примером колебаний предельного цикла, в котором частота ударов широко варьируется, в то время как каждое индивидуальное сердцебиение продолжает перекачивать примерно такое же количество крови.
Вычислительные модели используют множество абстракций для описания сложной колебательной динамики, наблюдаемой в активности мозга. В этой области используется множество моделей, каждая из которых определяется на разном уровне абстракции и пытается моделировать различные аспекты нейронных систем. Они варьируются от моделей краткосрочного поведения отдельных нейронов до моделей того, как динамика нейронных схем возникает из взаимодействий между отдельными нейронами, до моделей того, как поведение может возникать из абстрактных нейронных модулей, представляющих полные подсистемы.
 Имитация нейрона Хиндмарша – Роуза, демонстрирующая типичное взрывное поведение: быстрый ритм, генерируемый отдельными импульсами, и более медленный ритм, генерируемый всплески.
Имитация нейрона Хиндмарша – Роуза, демонстрирующая типичное взрывное поведение: быстрый ритм, генерируемый отдельными импульсами, и более медленный ритм, генерируемый всплески. Модель биологического нейрона - это математическое описание свойств нервных клеток или нейронов, предназначенное для точного описания и прогнозирования его биологических процессов. Наиболее успешная и широко используемая модель нейронов, модель Ходжкина – Хаксли, основана на данных, полученных с гигантского аксона кальмара. Это набор нелинейных обыкновенных дифференциальных уравнений, который аппроксимирует электрические характеристики нейрона, в частности генерацию и распространение потенциалов действия. Модель очень точна и детализирована, и Ходжкин и Хаксли получили Нобелевскую премию 1963 года по физиологии и медицине за эту работу.
Математика модели Ходжкина-Хаксли довольно сложна, и было предложено несколько упрощений, таких как модель ФитцХью-Нагумо, модель Хиндмарша-Роуза или модель конденсаторного переключателя как расширение модели интегрированного и пожарного. Такие модели отражают только базовую динамику нейронов, такую как ритмические всплески и взрыв, но они более эффективны с точки зрения вычислений. Это позволяет моделировать большое количество взаимосвязанных нейронов, которые образуют нейронную сеть.
Модель нейронной сети описывает совокупность физически связанных нейронов или группу разрозненных нейронов, входы которых или цели сигнализации определяют узнаваемую цепь. Эти модели призваны описать, как динамика нейронных схем возникает из взаимодействий между отдельными нейронами. Локальные взаимодействия между нейронами могут привести к синхронизации пиковой активности и сформировать основу осцилляторной активности. В частности, было показано, что модели взаимодействующих пирамидных клеток и тормозных интернейронов генерируют ритмы мозга, такие как гамма-активность. Аналогичным образом было показано, что моделирование нейронных сетей с феноменологической моделью отказов нейронного ответа может предсказать спонтанные широкополосные нейронные колебания.
 Моделирование модели нейронной массы, показывающее скачки сети в начале захват. По мере увеличения коэффициента усиления A сеть начинает колебаться с частотой 3 Гц.
Моделирование модели нейронной массы, показывающее скачки сети в начале захват. По мере увеличения коэффициента усиления A сеть начинает колебаться с частотой 3 Гц. Модели нейронного поля - еще один важный инструмент в изучении нейронных колебаний и математическая структура, описывающая эволюцию переменных, таких как средняя скорость возбуждения в пространстве и времени. При моделировании активности большого количества нейронов основная идея состоит в том, чтобы довести плотность нейронов до предела континуума, что приведет к пространственно непрерывным нейронным сетям. Вместо моделирования отдельных нейронов этот подход приближает группу нейронов по ее средним свойствам и взаимодействиям. Он основан на подходе среднего поля, области статистической физики, которая имеет дело с крупномасштабными системами. Модели, основанные на этих принципах, использовались для математического описания нервных колебаний и ритмов ЭЭГ. Например, их использовали для исследования зрительных галлюцинаций.
 Воспроизведение мультимедиа Имитация модели Курамото, демонстрирующая нейронную синхронизацию и колебания в среднем поле
Воспроизведение мультимедиа Имитация модели Курамото, демонстрирующая нейронную синхронизацию и колебания в среднем поле Модель Курамото связанных фазовых осцилляторов - одна из самых абстрактных и фундаментальных моделей, используемых для исследования нейронных колебаний и синхронизации. Он фиксирует активность локальной системы (например, одиночного нейрона или нейронного ансамбля) только с помощью своей круговой фазы и, следовательно, игнорирует амплитуду колебаний (амплитуда постоянна). Взаимодействия между этими осцилляторами вводятся простой алгебраической формой (такой как функция синус ) и вместе генерируют динамический образец в глобальном масштабе. Модель Курамото широко используется для изучения осцилляторной активности мозга, и было предложено несколько расширений, повышающих ее нейробиологическую достоверность, например, путем включения топологических свойств локальной корковой связности. В частности, он описывает, как активность группы взаимодействующих нейронов может синхронизироваться и генерировать крупномасштабные колебания. Моделирование с использованием модели Курамото с реалистичными корковыми связями на больших расстояниях и взаимодействиями с задержкой по времени показывает появление медленных шаблонных колебаний, которые воспроизводят функциональные карты состояния покоя ЖИРНЫЙ, которые можно измерить с помощью фМРТ.
И отдельные нейроны, и группы нейронов могут спонтанно генерировать колебательную активность. Кроме того, они могут показывать колебательные реакции на воспринимаемый вход или моторный выход. Некоторые типы нейронов будут срабатывать ритмично при отсутствии синаптического входа. Точно так же активность всего мозга выявляет колебательную активность, в то время как субъекты не участвуют в какой-либо деятельности, так называемая активность в состоянии покоя. Эти продолжающиеся ритмы могут изменяться по-разному в ответ на входные данные или моторные сигналы. Колебательная активность может реагировать увеличением или уменьшением частоты и амплитуды или показывать временное прерывание, которое называется сбросом фазы. Кроме того, внешняя активность может вообще не взаимодействовать с текущей активностью, что приводит к аддитивной реакции.

частота текущей колебательной активности увеличивается между t1 и t2.

амплитуда текущей колебательной активности увеличивается между t1 и t2.

Фаза текущей колебательной активности сбрасывается в t1.

Активность линейно добавляется к продолжающейся колебательной активности между t1 и t2.
Спонтанная активность - это активность мозга при отсутствии явной задачи, такой как сенсорный ввод или двигательный выход, и поэтому также называется активностью в состоянии покоя. Он противоположен индуцированной активности, то есть активности мозга, вызванной сенсорными стимулами или двигательными реакциями. Термин «текущая мозговая активность» используется в электроэнцефалографии и магнитоэнцефалографии для тех компонентов сигнала, которые не связаны с обработкой стимула или возникновением определенных других событий., например, перемещение части тела, т.е. события, которые не образуют вызванных потенциалов, / вызванных полей или индуцированной активности. Спонтанная активность обычно рассматривается как шум, если кто-то интересуется обработкой стимула; однако считается, что спонтанная активность играет решающую роль во время развития мозга, например, в формировании сети и синаптогенезе. Спонтанная активность может быть информативной относительно текущего психического состояния человека (например, бодрствования, бдительности) и часто используется в исследованиях сна. Определенные типы колебательной активности, такие как альфа-волны, являются частью спонтанной активности. Статистический анализ флуктуаций мощности альфа-активности показывает бимодальное распределение, то есть режим высокой и низкой амплитуды, и, следовательно, показывает, что активность в состоянии покоя не просто отражает процесс шум. В случае фМРТ спонтанные колебания сигнала , зависимого от уровня кислорода в крови (ЖИРНЫЙ), выявляют закономерности корреляции, которые связаны с сетями состояний покоя, такими как сеть по умолчанию. Временная эволюция сетей состояния покоя коррелирует с колебаниями осцилляторной активности ЭЭГ в разных частотных диапазонах.
Постоянная активность мозга также может играть важную роль в восприятии, поскольку она может взаимодействовать с активностью, связанной с поступающими стимулами. Действительно, исследования ЭЭГ предполагают, что зрительное восприятие зависит как от фазы, так и от амплитуды корковых колебаний. Например, амплитуда и фаза альфа-активности в момент зрительной стимуляции предсказывают, будет ли субъект воспринимать слабый стимул.
В ответ на ввод, нейрон или ансамбль нейронов может изменять частоту, с которой он колеблется, тем самым изменяя скорость, с которой он колеблется. Часто скорость активации нейрона зависит от суммарной активности, которую он получает. Частотные изменения также обычно наблюдаются в центральных генераторах паттернов и напрямую связаны со скоростью двигательной активности, например, с частотой шагов при ходьбе. Однако изменения относительной частоты колебаний между различными областями мозга не так распространены, потому что частота колебательной активности часто связана с временными задержками между областями мозга.
Помимо вызванной активности, нейронная активность, связанная с обработкой стимула, может привести к индуцированной активности. Индуцированная активность относится к модуляции текущей мозговой активности, вызванной обработкой стимулов или подготовкой к движению. Следовательно, они отражают косвенный ответ в отличие от вызванного ответа. Хорошо изученным видом индуцированной активности является изменение амплитуды колебательной активности. Например, гамма-активность часто увеличивается во время повышенной умственной активности, например, во время представления объекта. Поскольку индуцированные отклики могут иметь разные фазы при измерениях и, следовательно, отменяются при усреднении, их можно получить только с помощью частотно-временного анализа. Индуцированная активность обычно отражает активность множества нейронов: считается, что изменения амплитуды осцилляторной активности возникают из-за синхронизации нейронной активности, например, за счет синхронизации времени спайков или колебаний мембранного потенциала отдельных нейронов. Поэтому увеличение осцилляторной активности часто называют синхронизацией, связанной с событием, а уменьшение - десинхронизацией, связанной с событием.
Сброс фазы происходит при вводе в нейрон или нейрон ансамбль сбрасывает фазу идущих колебаний. Это очень часто встречается в отдельных нейронах, где синхронизация спайков регулируется в соответствии с входом нейрона (нейрон может давать всплеск с фиксированной задержкой в ответ на периодический вход, что называется фазовой синхронизацией), а также может возникать в ансамблях нейронов, когда фазы их нейроны настраиваются одновременно. Сброс фазы является фундаментальным для синхронизации разных нейронов или разных областей мозга, потому что время всплесков может стать синхронизированным по фазе с активностью других нейронов.
Сброс фазы также позволяет изучать вызванную активность - термин, используемый в электроэнцефалографии и магнитоэнцефалографии для ответов мозговой активности, которые напрямую связаны с стимулом деятельность, связанная с этим. Вызванные потенциалы и связанные с событием потенциалы получаются из электроэнцефалограммы путем усреднения с синхронизацией стимула, то есть усреднения различных испытаний с фиксированными латентными периодами вокруг предъявления стимула. Как следствие, те компоненты сигнала, которые одинаковы в каждом отдельном измерении, сохраняются, а все остальные, то есть текущая или спонтанная активность, усредняются. То есть потенциалы, связанные с событием, отражают только колебания мозговой активности, которые фаза привязаны к стимулу или событию. Вызванная активность часто считается независимой от текущей мозговой активности, хотя это продолжающиеся споры.
Недавно было предложено, что даже если фазы не совпадают в испытаниях, индуцированная активность может по-прежнему вызывать связанные с событием потенциалы, потому что текущие колебания мозга могут быть несимметричными, и, таким образом, амплитудные модуляции могут привести к смещению базовой линии, которое не усредняется. Эта модель предполагает, что медленные реакции, связанные с событиями, такие как асимметричная альфа-активность, могут быть результатом асимметричных модуляций амплитуды колебаний мозга, таких как асимметрия внутриклеточных токов, которые распространяются вперед и назад вниз по дендритам. Согласно этому предположению, асимметрия дендритного тока вызовет асимметрию осцилляторной активности, измеряемой с помощью ЭЭГ и МЭГ, поскольку обычно считается, что дендритные токи в пирамидных клетках генерируют сигналы ЭЭГ и МЭГ, которые можно измерить на коже черепа.
Нейронная синхронизация может модулироваться ограничениями задачи, такими как внимание, и считается, что она играет роль в привязке функций, нейронной коммуникации и координация движений. Нейрональные колебания стали горячей темой в нейробиологии в 1990-х годах, когда исследования зрительной системы мозга, проведенные Греем, Сингером и другими, подтвердили гипотезу нейронной привязки. Согласно этой идее, синхронные колебания в нейронных ансамблях связывают нейроны, представляющие различные характеристики объекта. Например, когда человек смотрит на дерево, нейроны зрительной коры, представляющие ствол дерева, и нейроны, представляющие ветви того же дерева, будут синхронно колебаться, чтобы сформировать единое представление дерева. Это явление лучше всего видно в потенциалах локального поля, которые отражают синхронную активность локальных групп нейронов, но также было показано в записях ЭЭГ и MEG, обеспечивающих увеличение доказательства тесной связи между синхронной осцилляторной активностью и различными когнитивными функциями, такими как перцептивная группировка.
Клетки в синоатриальном узле, расположенные в правое предсердие сердца, спонтанно деполяризуется примерно 100 раз в минуту. Хотя все клетки сердца обладают способностью генерировать потенциалы действия, которые запускают сердечное сокращение, синоатриальный узел обычно инициирует его просто потому, что он генерирует импульсы немного быстрее, чем другие области. Следовательно, эти клетки генерируют нормальный синусовый ритм и называются клетками кардиостимулятора, поскольку они непосредственно контролируют частоту сердечных сокращений. В отсутствие внешнего нейронального и гормонального контроля клетки в узле SA будут ритмично разряжаться. Синоатриальный узел обильно иннервируется вегетативной нервной системой, которая вверх или вниз регулирует частоту спонтанной активации пейсмекерных клеток.
Синхронизированное срабатывание нейронов также является основой периодических двигательных команд для ритмичных движений. Эти ритмические выходы производятся группой взаимодействующих нейронов, образующих сеть, называемую центральным генератором паттернов. Генераторы центральных паттернов представляют собой нейронные цепи, которые при активации могут создавать ритмические двигательные паттерны в отсутствие сенсорных или нисходящих входных сигналов, которые несут определенную временную информацию. Примеры: ходьба, дыхание и плавание. Большинство свидетельств генераторов центральных паттернов поступает от низших животных, таких как минога, но есть также свидетельства наличия генераторов центральных паттернов спинного мозга у людей.
Нейрональные импульсы обычно считаются основой для передачи информации в головном мозге. Для такой передачи информация должна быть закодирована в виде пиков. Были предложены различные типы схем кодирования, такие как кодирование со скоростью и временное кодирование. Нейронные колебания могут создавать периодические временные окна, в которых входные импульсы имеют большее влияние на нейроны, тем самым обеспечивая механизм для декодирования временных кодов.
Синхронизация возбуждения нейронов может служить средством группировки пространственно разделенные нейроны, которые реагируют на один и тот же стимул, чтобы связать эти ответы для дальнейшей совместной обработки, то есть использовать временную синхронность для кодирования отношений. Первыми были предложены чисто теоретические формулировки гипотезы связывания посредством синхронизации, но впоследствии появились обширные экспериментальные данные, подтверждающие потенциальную роль синхронности как кода отношений.
Функциональная роль синхронизированной колебательной активности в мозге в основном была установлена в экспериментах, проведенных на бодрствующих котятах с множественными электродами, имплантированными в зрительную кору. Эти эксперименты показали, что группы пространственно разделенных нейронов участвуют в синхронной осцилляторной активности при активации визуальными стимулами. Частота этих колебаний находилась в диапазоне 40 Гц и отличалась от периодической активации, вызванной решеткой, предполагая, что колебания и их синхронизация были вызваны внутренними взаимодействиями нейронов. Параллельно схожие находки были получены группой Eckhorn, предоставив дополнительные доказательства функциональной роли нейронной синхронизации в связывании признаков. С тех пор многочисленные исследования воспроизвели эти результаты и распространили их на различные методы, такие как ЭЭГ, предоставив обширные доказательства функциональной роли колебаний гамма в зрительном восприятии.
Жиль Лоран и его коллеги показали, что колебательная синхронизация играет важную функциональную роль в восприятии запахов. Восприятие разных запахов приводит к тому, что разные подмножества нейронов активируются в разных наборах колебательных циклов. Эти колебания могут быть нарушены ГАМК блокатором пикротоксином, и нарушение осцилляторной синхронизации приводит к нарушению поведенческой дискриминации химически схожих запахов у пчел и к более схожим ответам на запахи ниже по течению. β-долевые нейроны. Недавнее продолжение этой работы показало, что колебания создают периодические окна интегрирования для клеток Кеньона в теле насекомого грибовидное тело, так что входящие шипы из лепестка антенны более эффективны в активации клеток Кеньона только в определенных фазах колебательного цикла.
Считается, что нервные колебания также участвуют в чувстве времени и в соматосенсорном восприятии. Тем не менее, недавние открытия свидетельствуют против часовой функции кортикальных гамма-колебаний.
В двигательной системе часто наблюдаются колебания. Пфурчеллер и его коллеги обнаружили уменьшение колебаний альфа (8–12 Гц) и бета (13–30 Гц) в активности ЭЭГ, когда испытуемые совершали движение. Используя внутрикортикальные записи, аналогичные изменения колебательной активности были обнаружены в моторной коре, когда обезьяны выполняли двигательные действия, требующие значительного внимания. Кроме того, колебания на уровне позвоночника синхронизируются с бета-колебаниями в моторной коре при постоянной активации мышц, что определяется с помощью кортико-мышечной когерентности. Точно так же мышечная активность различных мышц обнаруживает межмышечную когерентность на нескольких различных частотах, отражающих лежащие в основе нейронные цепи, участвующие в координации движений.
. распространяются в виде бегущих волн по поверхности моторной коры по доминирующим пространственным осям, характерным для локальной схемы моторной коры. Было предложено, что двигательные команды в форме бегущих волн могут пространственно фильтроваться нисходящими волокнами для выборочного управления мышечной силой. Моделирование показало, что текущая волновая активность в коре головного мозга может вызывать устойчивую мышечную силу с физиологическими уровнями когерентности ЭЭГ-ЭМГ.
Колебательные ритмы с частотой 10 Гц были зарегистрированы в области мозга, называемой нижней оливой, который связан с мозжечком. Эти колебания также наблюдаются в моторном выходе физиологического тремора и при выполнении медленных движений пальцев. Эти данные могут указывать на то, что человеческий мозг периодически контролирует непрерывные движения. В подтверждение этого было показано, что эти нарушения непрерывности движений напрямую связаны с колебательной активностью в церебелло-таламо-кортикальной петле, которая может представлять нервный механизм для прерывистого моторного контроля.
Нервные колебания, в частности, активность тета, во многом связаны с функцией памяти. Тета-ритмы очень сильны в гиппокампе и энторинальной коре грызунов во время обучения и восстановления памяти, и они считаются жизненно важными для индукции долгосрочной потенциации, потенциального клеточного механизма обучения и памяти. Связь между тета и гамма активностью считается жизненно важным для функций памяти, включая эпизодическую память. Тесная координация спайков одиночных нейронов с локальными тета-колебаниями связана с успешным формированием памяти у людей, поскольку более стереотипные спайки предсказывают лучшую память.
Сон - это естественно повторяющееся состояние, характеризующееся сниженным или отсутствующим сознанием и продолжается циклами быстрого движения глаз (REM) и небыстрого движения глаз (NREM) сна. Стадии сна характеризуются спектральным содержанием ЭЭГ : например, стадия N1 относится к переходу мозга от альфа-волн (обычно в состоянии бодрствования) к тета-волнам, тогда как стадия N3 (глубокая или медленная- волновой сон) характеризуется наличием дельта-волн. Нормальный порядок стадий сна: N1 → N2 → N3 → N2 → REM.
Нервные колебания могут играть роль в нервном развитии. Например, считается, что волны на сетчатке обладают свойствами, которые определяют раннюю связь цепей и синапсов между клетками сетчатки.
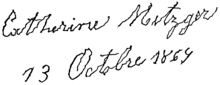 Почерк человека, страдающего болезнью Паркинсона. заболевание, показывающее ритмическую треморную активность при ударах
Почерк человека, страдающего болезнью Паркинсона. заболевание, показывающее ритмическую треморную активность при ударах  Общие спайки 3 Гц и волновые разряды, отражающие приступ активность
Общие спайки 3 Гц и волновые разряды, отражающие приступ активность Определенные типы нервных колебаний могут также возникать в патологических ситуациях, например Болезнь Паркинсона или эпилепсия. Эти патологические колебания часто представляют собой аберрантную версию нормальных колебаний. Например, один из наиболее известных типов - это спайковые и волновые колебания , которые типичны для генерализованных эпилептических припадков или абсансов и напоминают нормальные колебания веретена во сне.
Тремор - это непроизвольное, отчасти ритмичное мышечное сокращение и расслабление, включающее возвратно-поступательные движения одной или нескольких частей тела. Это наиболее распространенное из всех непроизвольных движений, которое может поражать руки, руки, глаза, лицо, голову, голосовые связки, туловище и ноги. Чаще всего тремор возникает в руках. У некоторых людей тремор является симптомом другого неврологического расстройства. Было идентифицировано множество различных форм тремора, таких как эссенциальный тремор или паркинсонический тремор. Утверждается, что тремор, вероятно, имеет многофакторное происхождение, в которое вносят вклад нервные колебания в центральной нервной системе, а также периферические механизмы, такие как резонансы рефлекторных петель.
Эпилепсия - это тоже обычное хроническое неврологическое заболевание, характеризующееся припадками. Эти припадки являются преходящими признаками и / или симптомами аномальной, чрезмерной или гиперсинхронной нейрональной активности в головном мозге.
При таламокортикальной дисритмии (TCD) нормальная таламокортикальный резонанс нарушен. Таламическая потеря входных данных позволяет частоте таламо-кортикального столбца замедляться до тета- или дельта-диапазона, что определяется с помощью MEG и EEG с помощью машинного обучения. TCD можно лечить с помощью нейрохирургических методов, таких как таламотомия.
Нервные колебания чувствительны к нескольким лекарствам, влияющим на активность мозга; соответственно, биомаркеры, основанные на нервных колебаниях, становятся вторичными конечными точками в клинических испытаниях и при количественной оценке эффектов в доклинических исследованиях. Эти биомаркеры часто называют «биомаркерами ЭЭГ» или «нейрофизиологическими биомаркерами» и количественно определяют с помощью количественной электроэнцефалографии (кЭЭГ). Биомаркеры ЭЭГ могут быть извлечены из ЭЭГ с использованием открытого источника Neurophysiological Biomarker Toolbox.
Нейронные колебания использовались в качестве управляющего сигнала в различных интерфейсах мозг – компьютер (BCI). Например, неинвазивный ИМК можно создать, поместив электроды на кожу головы и затем измерив слабые электрические сигналы. Хотя активность отдельных нейронов не может быть записана с помощью неинвазивного ИМК, поскольку череп демпфирует и размывает электромагнитные сигналы, колебательную активность все же можно надежно обнаружить. BCI был введен Видалем в 1973 году как задача использования сигналов ЭЭГ для управления объектами вне человеческого тела.
После испытания BCI в 1988 году альфа-ритм был использован в BCI, основанном на ритме мозга, для управления физическим объектом, роботом. BCI на основе альфа-ритма был первым BCI для управления роботом. В частности, некоторые формы BCI позволяют пользователям управлять устройством путем измерения амплитуды колебательной активности в определенных частотных диапазонах, включая mu и бета ритмы.
Неполный список типов колебательной активности, обнаруженных в центральной нервной системе: