
Войти
| Кора головного мозга | |
|---|---|
 Срез ткани мозга взрослой макаки-обезьяны (Macaca mulatta). Кора головного мозга - это внешний слой, изображенный темно-фиолетовым цветом. Источник: BrainMaps.org Срез ткани мозга взрослой макаки-обезьяны (Macaca mulatta). Кора головного мозга - это внешний слой, изображенный темно-фиолетовым цветом. Источник: BrainMaps.org | |
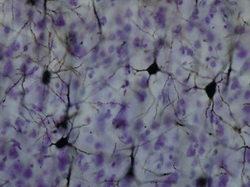 окрашенные по Гольджи нейроны в коре окрашенные по Гольджи нейроны в коре | |
| Подробности | |
| Часть | Cerebrum |
| Идентификаторы | |
| Латинские | Cortex cerebri |
| MeSH | D002540 |
| NeuroNames | 39 |
| NeuroLex ID | birnlex_1494 |
| TA98 | A14.1.09.003. A14.1.09.301 |
| TA2 | 5527, 5528 |
| FMA | 61830 |
| Анатомические термины нейроанатомии [редактировать в Викиданных ] | |
кора головного мозга (множественное число коры ), также известная как церебральная мантия, это внешний слой нервной ткани головного мозга мозга у людей и других млекопитающие. Кора головного мозга в основном состоит из шести слоев неокортекса, и только десять процентов состоит из аллокортекса. Он разделен на две коры продольной щелью, которая разделяет головной мозг на левое и правое полушария головного мозга. Два полушария под корой соединены мозолистым телом. Кора головного мозга является крупнейшим участком нейронной интеграции в центральной нервной системе. Он играет ключевую роль в внимании, восприятии, осведомленности, мысли, памяти, языке. и сознание.
У большинства млекопитающих, за исключением мелких млекопитающих с маленьким мозгом, кора головного мозга сложена, обеспечивая большую площадь поверхности в ограниченном объеме черепа. Помимо минимизации объема мозга и черепа, кортикальная складка имеет решающее значение для проводки мозга и его функциональной организации. У млекопитающих с маленьким мозгом складка отсутствует, а кора гладкая.
Складка или гребень в коре называется извилиной (множественные извилины), а бороздка называется борозда (множественное число борозд). Эти поверхностные извилины появляются во время развития плода и продолжают созревать после рождения в процессе гирификации. В человеческом мозге большая часть коры головного мозга не видна снаружи, но скрыта в бороздах, а островковая кора полностью скрыта. Основные борозды и извилины отмечают подразделения головного мозга на доли головного мозга.
В коре головного мозга имеется от 14 до 16 миллиардов нейронов. Они организованы в горизонтальные слои и радиально в кортикальные столбцы и мини-столбцы. Области коры имеют определенные функции, такие как движение в моторной коре и зрение в зрительной коре.
 Боковой вид коры
Боковой вид коры Кора головного мозга Внешнее покрытие поверхностей полушарий головного мозга состоит из пиков, называемых извилинами, и бороздок, называемых бороздами. В человеческом мозге он имеет толщину от двух до трех или четырех миллиметров и составляет 40 процентов массы мозга. 90 процентов коры головного мозга представляет собой шестислойный неокортекс, а остальные 10 процентов составляют аллокортекс. В коре имеется от 14 до 16 миллиардов нейронов, и они расположены радиально в кортикальных столбцах и мини-столбцах в горизонтально организованных слоях коры.
Около двух третей кортикальной поверхности скрыто в бороздах, а островковая кора полностью скрыта. Кора головного мозга наиболее толстая над вершиной извилины и самая тонкая в нижней части борозды.
Кора головного мозга сложена таким образом, чтобы обеспечить большую площадь поверхности нервная ткань, чтобы уместиться в пределах нейрокраниума. В развернутом виде у человека каждая полусферическая кора головного мозга имеет общую площадь поверхности около 0,12 квадратных метра (1,3 квадратных фута). Складка направлена внутрь от поверхности мозга, а также присутствует на медиальной поверхности каждого полушария в пределах продольной щели. У большинства млекопитающих кора головного мозга извилина с пиками, известными как извилины, и впадинами или бороздками, известными как борозды. У некоторых мелких млекопитающих, включая некоторых мелких грызунов, церебральные поверхности гладкие без гирификации.
Более крупные борозды и извилины отмечают подразделения коры головного мозга на доли головного мозга. Всего четыре основные доли: лобная доля, теменная доля, височная доля и затылочная доля. кора островка часто включается как островная доля. лимбическая доля - это край коры на медиальной стороне каждого полушария, который также часто включается. Также описаны три доли головного мозга: парацентральная доля, верхняя теменная долька и нижняя теменная доля.
Для У видов млекопитающих более крупный мозг (в абсолютном выражении, а не только по отношению к размеру тела), как правило, имеет более толстую корку. Самые маленькие млекопитающие, такие как землеройки, имеют толщину неокортекса около 0,5 мм; те, у кого самый большой мозг, такие как люди и финвалы, имеют толщину 2–4 мм. Существует приблизительно логарифмическое соотношение между массой мозга и толщиной коры. Магнитно-резонансная томография головного мозга (МРТ) позволяет измерить толщину коры головного мозга человека и соотнесите это с другими мерами. Толщина различных областей коры различается, но в целом сенсорная кора тоньше моторной коры. Одно исследование обнаружило некоторую положительную связь между толщиной коры и интеллектом. Другое исследование показало, что соматосенсорная кора толще у людей, страдающих мигренью,, хотя неизвестно, является ли это результатом приступов мигрени или их причиной. В более позднем исследовании с участием большего количества пациентов сообщается об отсутствии изменений в толщине коркового слоя у страдающих мигренью. Генетическое заболевание коры головного мозга, при котором уменьшение складчатости в определенных областях приводит к микрогирусу, где четыре слоя вместо шести, в некоторых случаях связано с дислексией.
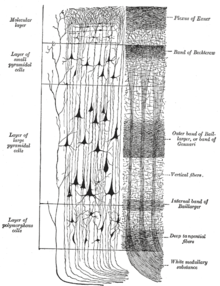 Кора головного мозга. (Пуарье.) Слева - группы ячеек; справа - системы волокон. Слева от рисунка показано чувствительное нервное волокно. Слои клеточного тела обозначены слева, а слои волокон - справа.
Кора головного мозга. (Пуарье.) Слева - группы ячеек; справа - системы волокон. Слева от рисунка показано чувствительное нервное волокно. Слои клеточного тела обозначены слева, а слои волокон - справа. 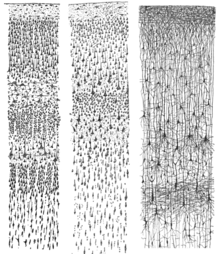 Три рисунка кортикального ламинирования, выполненные Сантьяго Рамон-и-Кахал, каждый из которых показывает вертикальное поперечное сечение с поверхностью кора наверху. Слева: окрашенная по Нисслю зрительная кора взрослого человека. В центре: окрашенная по Нисслю моторная кора взрослого человека. Справа: -окрашенная по Гольджи кора головного мозга ребенка в возрасте 1 / 2 месяца. Окрашивание по Нисслю показывает клеточные тела нейронов; окрашивание по Гольджи показывает дендриты и аксоны случайного подмножества нейронов.
Три рисунка кортикального ламинирования, выполненные Сантьяго Рамон-и-Кахал, каждый из которых показывает вертикальное поперечное сечение с поверхностью кора наверху. Слева: окрашенная по Нисслю зрительная кора взрослого человека. В центре: окрашенная по Нисслю моторная кора взрослого человека. Справа: -окрашенная по Гольджи кора головного мозга ребенка в возрасте 1 / 2 месяца. Окрашивание по Нисслю показывает клеточные тела нейронов; окрашивание по Гольджи показывает дендриты и аксоны случайного подмножества нейронов.  Микрофотография, показывающая зрительную кору (преимущественно розовую). Подкорковое белое вещество (преимущественно синее) видно в нижней части изображения. Окраска HE-LFB.
Микрофотография, показывающая зрительную кору (преимущественно розовую). Подкорковое белое вещество (преимущественно синее) видно в нижней части изображения. Окраска HE-LFB.неокортекс состоит из шести кортикальных слоев, пронумерованных от I до VI от самого внешнего (прилегающего к мягкой мозговой оболочке ) до самого внутреннего (прилегающего в белое вещество ). Каждый слой имеет характерное распределение разных нейронов и их связи с другими корковыми и подкорковыми областями. Есть прямые связи между различными областями коры и косвенные связи через таламус.
Одним из наиболее ярких примеров кортикального наслоения является линия Геннари в первичной зрительной коре. Это полоса более белой ткани, которую можно наблюдать невооруженным глазом на дне калькариновой борозды затылочной доли. Линия Геннари состоит из аксонов, доставляющих визуальную информацию из таламуса в слой IV зрительной коры.
Окрашивание поперечных срезов коры для Выявление положения тел нейронных клеток и интракортикальных трактов аксонов позволило нейроанатомам в начале 20 века произвести подробное описание ламинарной структуры коры у разных видов. В работе Корбиниана Бродмана (1909) установлено, что неокортекс млекопитающих (и большинство других областей коры) последовательно делится на шесть слоев.
Слой I является молекулярным слоем и содержит несколько разбросанных нейронов, включая ГАМКергические нейроны шиповника. Слой I состоит в основном из расширений апикальных дендритных пучков пирамидных нейронов и горизонтально ориентированных аксонов, а также глиальных клеток. Во время развития клетки Кахаля-Ретциуса и клетки субпиального гранулярного слоя присутствуют в этом слое. Также здесь можно найти несколько колючих звездчатых клеток. Считается, что входы в апикальные пучки имеют решающее значение для обратных взаимодействий в коре головного мозга, участвующих в ассоциативном обучении и внимании. Хотя когда-то считалось, что вход в слой I поступает из самой коры головного мозга, теперь стало понятно, что слой I, проходящий через мантию коры головного мозга, получает существенный вклад от клеток таламуса матрицы или M-типа (в отличие от ядер или клеток C-типа, которые перейти к слою IV).
Слой II, внешний зернистый слой, содержит небольшие пирамидные нейроны и многочисленные звездчатые нейроны.
Слой III, внешний пирамидный слой, содержит преимущественно пирамидные нейроны малого и среднего размера, а также непирамидные нейроны с вертикально ориентированными внутрикортикальными аксонами ; слои с I по III являются основной мишенью межполушарных кортикокортикальных афферентов, а слой III является основным источником кортикокортикальных эфферентов.
Слой IV, внутренний зернистый слой, содержит различные типы звездчатых и пирамидных клеток, и является основной мишенью таламокортикальных афферентов нейронов таламуса типа C (сердцевинного типа), а также внутри -полушарные корково-корковые афференты. Слои над слоем IV также называются супрагранулярными слоями (слои I-III), тогда как слои ниже называются инфрагранулярными слоями (слои V и VI).
Слой V, внутренний пирамидный слой, содержит большие пирамидальные нейроны. Аксоны из них покидают кору и соединяются с подкорковыми структурами, включая базальные ганглии. В первичной моторной коре лобной доли слой V содержит гигантские пирамидные клетки, называемые клетками Беца, аксоны которых проходят через внутреннюю капсулу, ствол мозга, и спинной мозг, образующий кортикоспинальный тракт, который является основным путем для произвольного моторного контроля.
Слой VI, полиморфный или многоформный слой, содержит несколько больших пирамидальных нейронов и множество маленьких веретенообразных пирамидных и многоформных нейронов; слой VI посылает эфферентные волокна в таламус, устанавливая очень точное взаимное соединение между корой и таламусом. То есть нейроны слоя VI из одного коркового столбца соединяются с нейронами таламуса, которые обеспечивают вход в тот же кортикальный столбец. Эти связи являются как возбуждающими, так и тормозящими. Нейроны посылают возбуждающие волокна к нейронам таламуса, а также посылают коллатерали в ретикулярное ядро таламуса, которое ингибирует эти же нейроны таламуса или соседние с ними. Одна из теорий заключается в том, что, поскольку тормозящий эффект снижается холинергическим входом в кору головного мозга, это обеспечивает ствол мозга регулируемым «контролем усиления для реле лемнискаля. входы ».
Кортикальные слои не просто накладываются друг на друга; существуют характерные связи между различными слоями и типами нейронов, охватывающими всю толщину коры. Эти корковые микросхемы сгруппированы в кортикальные столбцы и миниколонки. Было высказано предположение, что мини-столбцы являются основными функциональными единицами коры головного мозга. В 1957 г. Вернон Маунткасл показал, что функциональные свойства коры резко меняются между соседними латерально точками; однако они непрерывны в направлении, перпендикулярном поверхности. Более поздние работы предоставили доказательства присутствия функционально различных корковых столбов в зрительной коре (Hubel and Wiesel, 1959), слуховой коре и ассоциативной коре.
Области коры, в которых отсутствует слой IV, называются агранулярными. Области коры, которые имеют только рудиментарный слой IV, называются дисгранулярными. Обработка информации в каждом слое определяется разной временной динамикой: в слоях II / III наблюдается медленное колебание 2 Hz , а в слое V - быстрое колебание 10–15 Гц.
Основываясь на различиях в ламинарной организации, кору головного мозга можно разделить на два типа: большая область неокортекса, которая имеет шесть слоев клеток, и намного меньшая область аллокортекса, которая имеет три или четыре слоя:
. Между неокортексом и аллокортексом есть переходная область, называемая паралимбической корой, где слои 2, 3 и 4 объединены. Эта область включает произокортекс неокортекса и периаллокортекс аллокортекса. Кроме того, кору головного мозга можно разделить на четыре доли : лобную долю, височную долю, теменную долю и затылочная доля, названная в честь лежащих на них костей черепа.
 Артериальное кровоснабжение, показывающее области, снабжаемые задней, средней и передней церебральными артериями.
Артериальное кровоснабжение, показывающее области, снабжаемые задней, средней и передней церебральными артериями.Кровоснабжение коры головного мозга является частью мозгового кровообращения. Церебральные артерии снабжают кровью, кровоснабжающей головной мозг. Эта артериальная кровь переносит кислород, глюкозу и другие питательные вещества в кору. Церебральные вены отводят дезоксигенированную кровь и метаболические отходы, включая углекислый газ, обратно в сердце.
Основными артериями, снабжающими кору головного мозга, являются передняя церебральная артерия, средняя мозговая артерия и задняя мозговая артерия. Передняя мозговая артерия снабжает передние отделы головного мозга, включая большую часть лобной доли. Средняя мозговая артерия снабжает кровью теменные доли, височные доли и части затылочных долей. Средняя мозговая артерия разделяется на две ветви, чтобы снабжать левое и правое полушария, где они разветвляются дальше. Задняя мозговая артерия снабжает затылочные доли.
Уиллисовский круг - это основная система крови, которая отвечает за кровоснабжение головного мозга и коры головного мозга.
 Кортикальное кровоснабжение
Кортикальное кровоснабжение пренатальное развитие коры головного мозга - это сложный и точно настроенный процесс, называемый кортикогенез, на который влияет взаимодействие между гены и окружающая среда.
Кора головного мозга развивается из самой передней части, области переднего мозга, нервной трубки. нервная пластинка складывается и закрывается, образуя нервную трубку. Из полости внутри нервной трубки развивается желудочковая система, а из нейроэпителиальных клеток его стенок - нейроны и глия нервной системы. Самая передняя (передняя или краниальная) часть нервной пластинки, передний мозг, которая проявляется до начала нейруляции, дает начало полушариям головного мозга, а затем и коре.
Кортикальные нейроны генерируются в желудочковой зоне, рядом с желудочками. Сначала эта зона содержит нервные стволовые клетки, которые переходят в радиальные глиальные клетки - клетки-предшественники, которые делятся с образованием глиальных клеток и нейронов.
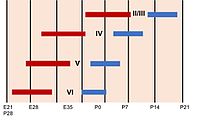 Нейрогенез показан красным цветом, а расслоение - синим. Взято из (Sur et al. 2001)
Нейрогенез показан красным цветом, а расслоение - синим. Взято из (Sur et al. 2001) Кора головного мозга состоит из гетерогенной популяции клеток, которые дают начало различным типам клеток. Большинство этих клеток происходит от миграции радиальной глии, которая формирует различные типы клеток неокортекса, и этот период связан с усилением нейрогенеза. Точно так же процесс нейрогенеза регулирует ламинирование с образованием различных слоев коры. Во время этого процесса происходит усиление ограничения судьбы клеток, которое начинается с более ранних предшественников, дающих начало любому типу клеток в коре головного мозга, и более поздних предшественников, дающих начало только нейронам поверхностных слоев.. Эта дифференцированная судьба клеток создает в коре головного мозга вывернутую топографию с более молодыми нейронами в поверхностных слоях и старыми нейронами в более глубоких слоях. Кроме того, ламинарные нейроны останавливаются в S или G2 фазе, чтобы дать четкое различие между разными корковыми слоями. Ламинарная дифференцировка не полностью завершается до момента рождения, поскольку во время развития ламинарные нейроны все еще чувствительны к внешним сигналам и сигналам окружающей среды.
Хотя большинство клеток, составляющих кору, происходят локально из радиальной глии, существует подмножество популяция нейронов, которая мигрирует из других регионов. Радиальная глия дает начало нейронам пирамидальной формы, использующим глутамат в качестве нейромедиатора, однако эти мигрирующие клетки вносят вклад в нейроны, имеющие звездчатую форму и использующие ГАМК как их главный нейротрансмиттер. Эти ГАМКергические нейроны генерируются клетками-предшественниками в медиальном ганглиозном возвышении (MGE), которые мигрируют тангенциально к коре через субвентрикулярную зону. Эта миграция ГАМКергических нейронов особенно важна, поскольку рецепторы ГАМК являются возбуждающими во время развития. Это возбуждение в основном вызывается потоком ионов хлора через рецептор ГАМК, однако у взрослых концентрация хлоридов смещается, вызывая входящий поток хлорида , который гиперполяризует постсинаптические нейроны. Глиальные волокна, образующиеся в первых отделах клеток-предшественников, ориентированы радиально, охватывая толщину коры от желудочковой зоны до внешней пиальной поверхности, и обеспечивают основу для миграция нейронов наружу из желудочковой зоны.
При рождении на теле клетки кортикального нейрона присутствует очень мало дендритов, а аксон не развит. В течение первого года жизни количество дендритов резко увеличивается, так что они могут вместить до ста тысяч синаптических связей с другими нейронами. Аксон может развиваться и простираться далеко от тела клетки.
Первые деления клеток-предшественников симметричны, что дублирует общее количество клеток-предшественников на каждом митотический цикл. Затем некоторые клетки-предшественники начинают делиться асимметрично, производя одну постмитотическую клетку, которая мигрирует по радиальным глиальным волокнам, покидая желудочковую зону, и одну клетку-предшественницу, которая продолжает делиться до конца развития, когда она дифференцируется в глиальную клетку или эпендимную клетку. Поскольку фаза G1 в митозе удлиняется, что рассматривается как избирательное удлинение клеточного цикла, новорожденные нейроны мигрируют в более поверхностные слои коры. Мигрирующие дочерние клетки становятся пирамидными клетками коры головного мозга. Процесс развития упорядочен и регулируется во времени сотнями генов и эпигенетическими регуляторными механизмами.
 Развитие коры мозга человека между 26 и 39 неделями гестации
Развитие коры мозга человека между 26 и 39 неделями гестации слоистая структура зрелая кора головного мозга формируется в процессе развития. Первые сгенерированные пирамидные нейроны мигрируют из желудочковой зоны и субвентрикулярной зоны вместе с продуцирующими рилин нейронами Кахаля-Ретциуса из предварительная пластина . Затем когорта нейронов, мигрирующих в середину препластинки, делит этот переходный слой на поверхностную маргинальную зону, которая станет слоем I зрелого неокортекса, и субпластинку, формируя средний слой, называемый корковой пластинкой . Эти клетки образуют глубокие слои зрелой коры, пятый и шестой слои. Позже рожденные нейроны мигрируют радиально в кортикальную пластинку, минуя нейроны глубокого слоя, и становятся верхними слоями (двумя-четырьмя). Таким образом, слои коры создаются в порядке «наизнанку». Единственное исключение из этой вывернутой наизнанку последовательности нейрогенеза встречается в слое I приматов, в котором, в отличие от грызунов, нейрогенез продолжается на протяжении всего период кортикогенеза.
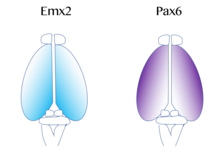 Изображенный синим цветом, Emx2 высоко экспрессируется на каудомедиальном полюсе и рассеивается наружу. Экспрессия Pax6 представлена пурпурным цветом и сильно выражена на боковом ростральном полюсе. (По материалам Sanes, D., Reh, T., Harris, W. (2012). Развитие нервной системы (3-е изд.). Burlington: Elsevier Science)
Изображенный синим цветом, Emx2 высоко экспрессируется на каудомедиальном полюсе и рассеивается наружу. Экспрессия Pax6 представлена пурпурным цветом и сильно выражена на боковом ростральном полюсе. (По материалам Sanes, D., Reh, T., Harris, W. (2012). Развитие нервной системы (3-е изд.). Burlington: Elsevier Science) Карта функциональных областей коры головного мозга, которые включают первичная моторная и зрительная кора происходит из «протокарты », которая регулируется молекулярными сигналами, такими как фактор роста фибробластов FGF8 на ранних стадиях эмбрионального развития. Эти сигналы регулируют размер, форму и положение корковых областей на поверхности кортикального зачатка, частично регулируя градиенты экспрессии фактора транскрипции, посредством процесса, называемого формированием коркового паттерна. Примеры таких факторов транскрипции включают гены EMX2 и PAX6. Вместе оба фактора транскрипции образуют противоположный градиент экспрессии. Pax6 высоко экспрессируется на ростральном латеральном полюсе, тогда как Emx2 высоко экспрессируется в каудомедиальном полюсе. Установление этого градиента важно для правильного развития. Например, мутации в Pax6 могут вызвать расширение уровней экспрессии Emx2 за пределы его нормального домена экспрессии, что в конечном итоге приведет к расширению областей, обычно происходящих из каудальной медиальной коры, таких как зрительная кора. Напротив, если возникают мутации в Emx2, это может вызвать расширение экспрессирующего Pax6 домена и привести к увеличению лобной и моторной кортикальной областей. Следовательно, исследователи полагают, что подобные градиенты и сигнальные центры рядом с корой могут способствовать региональной экспрессии этих факторов транскрипции. Два очень хорошо изученных паттернирующих сигнала для коры головного мозга включают FGF и ретиноевую кислоту. Если FGF неправильно экспрессируются в разных областях развивающейся коры, формирование коркового паттерна нарушается. В частности, когда Fgf8 увеличивается в переднем полюсе, Emx2 подавляется и происходит каудальный сдвиг в области коры. В конечном итоге это вызывает расширение ростральных областей. Следовательно, Fgf8 и другие FGF играют роль в регуляции экспрессии Emx2 и Pax6 и представляют, как кора головного мозга может специализироваться для различных функций.
Быстрое расширение площади поверхности коры регулируется количеством самообновление радиальных глиальных клеток и частично регулируется FGF и генами Notch. В период коркового нейрогенеза и формирования слоев у многих высших млекопитающих начинается процесс гирификации, который генерирует характерные складки коры головного мозга. Гирификация регулируется ДНК-ассоциированным белком Trnp1, а также FGF и SHH сигнализирующими
Из всех различных областей мозга церебральные кора головного мозга показывает самые большие эволюционные вариации и развивалась совсем недавно. В отличие от высококонсервативной схемы, например, продолговатого мозга, который выполняет важные функции, такие как регуляция частоты сердечных сокращений и дыхания, многие области коры головного мозга не являются строго необходимыми для выживания. Таким образом, эволюция коры головного мозга привела к появлению и модификации новых функциональных областей - в частности, ассоциативных областей, которые не получают непосредственного ввода извне коры.
Ключевая теория корковой эволюции воплощена в гипотеза радиальных единиц и связанная с ними гипотеза протокарты, впервые предложенная Ракичем. Эта теория утверждает, что новые области коры формируются путем добавления новых радиальных единиц, что достигается на уровне стволовых клеток. Гипотеза протокарты утверждает, что клеточная и молекулярная идентичность и характеристики нейронов в каждой области коры определяются кортикальными стволовыми клетками, известными как радиальные глиальные клетки, на первичной карте. Эта карта контролируется секретируемыми сигнальными белками и нижестоящими факторами транскрипции.
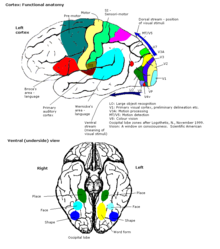 Некоторые функциональные области коры
Некоторые функциональные области коры Кора головного мозга связана с различными подкорковые структуры, такие как таламус и базальные ганглии, посылая им информацию по эфферентным связям и получая от них информацию через афферентные связи. Большая часть сенсорной информации направляется в кору головного мозга через таламус. Однако обонятельная информация проходит через обонятельную луковицу в обонятельную кору (грушевидная кора ). Большинство соединений идет от одной области коры к другой, а не от подкорковых областей; Braitenberg и Schüz (1998) утверждают, что в первичных сенсорных областях, на кортикальном уровне, где заканчиваются входные волокна, до 20% синапсов снабжается экстракортикальными афферентами, но в других областях и других слоях
 Боковая поверхность коры головного мозга человека
Боковая поверхность коры головного мозга человека  Медиальная поверхность коры головного мозга человека
Медиальная поверхность коры головного мозга человека Вся кора головного мозга была разделена на 52 различных области в раннем изложении Корбиняна Бродмана. Эти области, известные как области Бродмана, основаны на их цитоархитектуре, но также связаны с различными функциями. Примером является область 17 Бродмана, которая является первичной зрительной корой.
В более общем плане кора головного мозга обычно описывается как состоящая из трех частей: сенсорной, моторной и ассоциативной областей.
Сенсорные зоны - это области коры, которые получают и обрабатывают информацию от органов чувств. Части коры, которые получают сенсорные сигналы от таламуса, называются первичными сенсорными областями. Чувства зрения, слуха и осязания обслуживаются первичной зрительной корой, первичной слуховой корой и первичной соматосенсорной корой соответственно. В общем, два полушария получают информацию с противоположной (контралатеральной) стороны тела. Например, правая первичная соматосенсорная кора получает информацию от левых конечностей, а правая зрительная кора получает информацию от левого зрительного поля . Организация сенсорных карт в коре головного мозга отражает организацию соответствующего сенсорного органа на так называемой топографической карте. Соседние точки в первичной зрительной коре, например, соответствуют соседним точкам в сетчатке. Эта топографическая карта называется ретинотопической картой. Таким же образом существует тонотопическая карта в первичной слуховой коре и соматотопическая карта в первичной сенсорной коре. Эта последняя топографическая карта тела задней центральной извилины была проиллюстрирована как деформированное человеческое изображение, соматосенсорный гомункул, где размер различных частей тела отражает относительную плотность их иннервация. Области с высокой сенсорной иннервацией, такие как кончики пальцев и губы, требуют большей корковой области для обработки более тонких ощущений.
Двигательные области расположены в обоих полушариях коры. Двигательные области очень тесно связаны с контролем произвольных движений, особенно мелких фрагментированных движений, выполняемых рукой. Правая половина моторной области контролирует левую сторону тела, и наоборот.
Две области коры обычно называют моторными:
Кроме того, моторные функции описаны для:
Прямо под корой головного мозга находятся взаимосвязанные подкорковые массы серого вещества, называемые базальными ганглиями (или ядрами). Базальные ганглии получают сигнал от черной субстанции среднего мозга и моторных областей коры головного мозга и посылают сигналы обратно в оба этих места. Они участвуют в двигательном контроле. Они находятся сбоку от таламуса. Основными компонентами базальных ганглиев являются хвостатое ядро , скорлупа, бледный шар, черная субстанция, nucleus accumbens и субталамическое ядро . Скорлупа и бледный шар также вместе известны как лентиформное ядро , потому что вместе они образуют линзовидное тело. Скорлупа и хвостатое ядро также вместе называются полосатым телом после их полосатого вида.
 Области коры, участвующие в обработке речи.
Области коры, участвующие в обработке речи. Области ассоциации - части коры головного мозга, которые не относятся к первичным областям. Они функционируют для создания осмысленного перцептивного опыта мира, позволяют нам эффективно взаимодействовать и поддерживают абстрактное мышление и язык. теменная, височная и затылочная доли, расположенные в задней части коры головного мозга, объединяют сенсорную информацию и информацию, хранящуюся в памяти. лобная доля или комплекс префронтальных ассоциаций участвует в планировании действий и движений, а также в абстрактном мышлении. В глобальном масштабе зоны ассоциации организованы как распределенные сети. Каждая сеть соединяет области, распределенные по широко разнесенным областям коры. Отдельные сети расположены рядом друг с другом, образуя сложную серию переплетенных сетей. Конкретная организация ассоциативных сетей обсуждается с доказательствами взаимодействий, иерархических отношений и конкуренции между сетями.
У людей ассоциативные сети особенно важны для языковой функции. Ранее предполагалось, что языковые способности локализуются в области Брока в областях левой нижней лобной извилины, BA44 и BA45, для языкового выражения и в области Вернике BA22, для языкового приема. Однако было показано, что процессы языкового выражения и восприятия происходят не только в этих структурах вокруг боковой борозды, включая лобную долю, базальные ганглии, мозжечок. и pons.
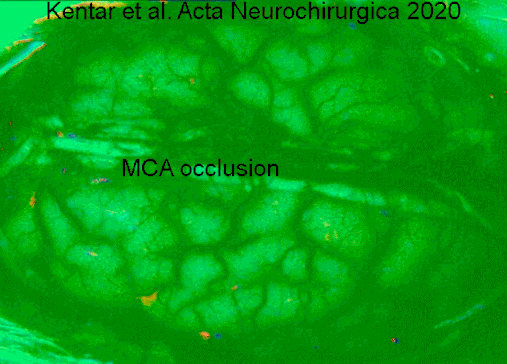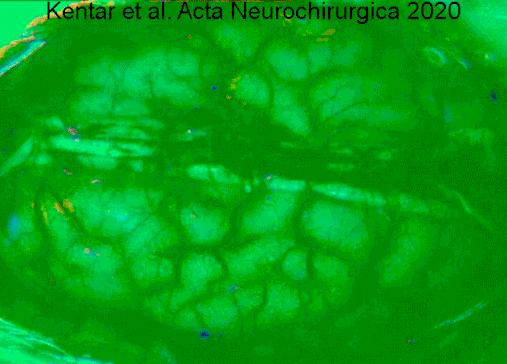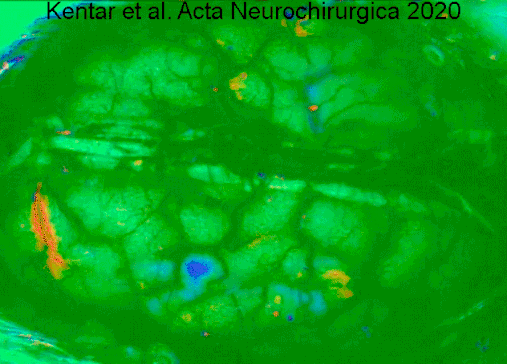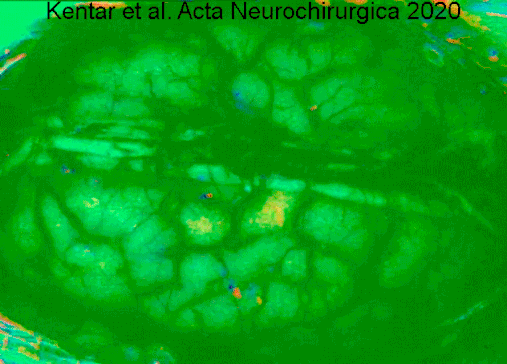 Гемодинамические изменения, наблюдаемые в коре головного мозга гринцефалической области после окклюзии артериального сосуда в IOS. Видео имеет скорость 50x, чтобы лучше оценить деполяризацию, распространяющуюся по коре головного мозга. Изображения динамически вычитают к опорному кадру 40 с раньше. Сначала мы видим первоначальные изменения в точный момент, когда группа средней мозговой артерии (слева) окклюзирована. Область выделена белой линией. Позже мы оценим сигнал, производимый расширяющейся деполяризацией. Мы отчетливо видим фронт волн. https://doi.org/10.1007/s00701-019-04132-8
Гемодинамические изменения, наблюдаемые в коре головного мозга гринцефалической области после окклюзии артериального сосуда в IOS. Видео имеет скорость 50x, чтобы лучше оценить деполяризацию, распространяющуюся по коре головного мозга. Изображения динамически вычитают к опорному кадру 40 с раньше. Сначала мы видим первоначальные изменения в точный момент, когда группа средней мозговой артерии (слева) окклюзирована. Область выделена белой линией. Позже мы оценим сигнал, производимый расширяющейся деполяризацией. Мы отчетливо видим фронт волн. https://doi.org/10.1007/s00701-019-04132-8Нейродегенеративные заболевания, такие как болезнь Альцгеймера и болезнь Лафора, указываются в качестве маркера, атрофия серое вещество коры головного мозга.
Другие заболевания центральной нервной системы включают неврологические расстройства, такие как эпилепсия, движение расстройства и затруднения речи (афазия ).
Повреждение мозга в результате болезни или травмы, может включать повреждение определенной доли, например, при заболевании лобной доли, и связанные с ним функции будут затронуты изд. гематоэнцефалический барьер, который защищает мозг от инфекции, может быть нарушен, что приведет к проникновению патогенов.
развивающийся плод восприимчив к ряду факторов окружающей среды, которые может вызвать врожденные дефекты и проблемы в дальнейшем развитии. Например, употребление алкоголя матерью может вызвать расстройство алкогольного спектра плода. Другими факторами, которые могут вызвать нарушения развития нервной системы, являются токсиканты, такие как лекарства, и воздействие радиации, например, рентгеновские лучи. Инфекции также могут повлиять на развитие коры головного мозга. Вирусная инфекция является одной из причин лиссэнцефалии, которая приводит к гладкой коре без гирификации.
Тип электрокортикографии, называемый картирование стимуляции коры - это инвазивная процедура, при которой электроды помещаются непосредственно на обнаженный мозг, чтобы локализовать функции определенных областей коры. Он используется в клинических и терапевтических целях, включая предоперационное картирование.
Существует ряд генетических мутаций, которые могут вызывать широкий спектр генетических нарушений коры головного мозга, включая микроцефалию, шизэнцефалию и типы лиссэнцефалии. Хромосомные аномалии также могут вызывать ряд расстройства нервного развития, такие как синдром ломкой Х-хромосомы и синдром Ретта.
MCPH1 коды для микроцефалина, а также нарушения в этом и в ASPM связаны с микроцефалией. Мутации в гене NBS1, который кодирует нибрин, могут вызывать синдром разрушения Неймегена, характеризующийся микроцефалией.
Мутации в EMX2 и COL4A1 связаны с шизэнцефалией, состоянием, при котором отсутствуют большие части полушарий головного мозга.
В 1909 году Корбиниан Бродманн выделил различные области неокортекса на основе цитоархитектурных различий и разделил кору головного мозга на 52 области.
Рафаэль Лоренте де Но, студент Сантьяго Рамон-и-Кахал идентифицировал более 40 различных типов корковых нейронов на основе распределения их дендритов и аксонов.
Кора головного мозга происходит от pallium, слоистой структуры, обнаруженной в переднем мозге всех позвоночных. Основная форма паллия - это цилиндрический слой, охватывающий желудочки, заполненные жидкостью. По окружности цилиндра расположены четыре зоны: дорсальный паллий, медиальный паллий, вентральный паллий и латеральный паллий, которые, как считается, соответственно дают начало неокортексу, гиппокампу, миндалевидное тело и обонятельная кора.
До недавнего времени не было обнаружено аналога коры головного мозга у беспозвоночных. Однако исследование, опубликованное в журнале Cell в 2010 году, основанное на профилях экспрессии генов, показало сильное сродство между корой головного мозга и грибовидными телами щупальца Platynereis dumerilii. Грибные тела - это структуры в мозге многих типов червей и членистоногих, которые, как известно, играют важную роль в обучении и памяти; генетические данные указывают на общее эволюционное происхождение и, следовательно, указывают на то, что происхождение самых ранних предшественников коры головного мозга восходит к ранней докембрийской эре.

Моторные и сенсорные области коры головного мозга

Моторные и сенсорные области коры головного мозга
{kind=link}