
Войти
| Невруляция | |
|---|---|
 Поперечные срезы, которые показывают продвижение нервной пластинки к нервной борозде от снизу вверх Поперечные срезы, которые показывают продвижение нервной пластинки к нервной борозде от снизу вверх | |
| Идентификаторы | |
| MeSH | D054261 |
| Анатомическая терминология [правка в Викиданных ] | |
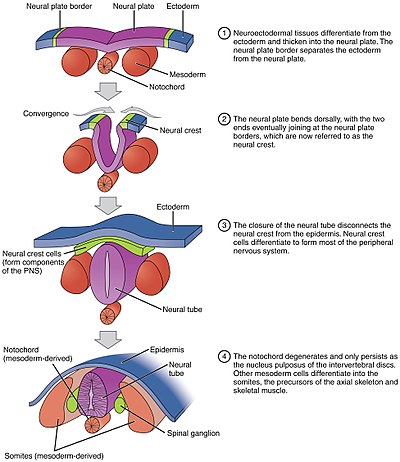
Невруляция относится к процессу складывания в эмбрионах позвоночных , который включает преобразование нервной пластинки в нервную трубку. Эмбрион на этой стадии называется нейрулой.
. Процесс начинается, когда хорда индуцирует образование центральной нервной системы (ЦНС), передавая сигналы эктодерме зародышевый слой над ним, образуя толстую и плоскую нервную пластинку. Нервная пластинка складывается сама по себе, образуя нервную трубку, которая позже дифференцируется на спинной мозг и головной мозг, в конечном итоге формируя центральную нервную систему. Компьютерное моделирование показало, что расклинивание клеток и дифференциальная пролиферация достаточны для нейруляции млекопитающих.
Различные части нервной трубки формируются двумя разными процессами, называемыми первичной и вторичной нейруляцией, у разных видов.
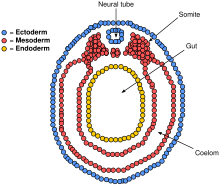 Поперечный разрез эмбриона позвоночного в стадия нейрулы
Поперечный разрез эмбриона позвоночного в стадия нейрулы  Воспроизвести медиа Описание процесса нейруляции в трех измерениях.
Воспроизвести медиа Описание процесса нейруляции в трех измерениях. Концепция индукции возникла в работе Пандор в 1817 году. Первые эксперименты, подтверждающие индукцию, Виктор Гамбургер приписал независимым открытиям как Ганса Спемана из Германии в 1901 году, так и Уоррена Льюиса из США в 1904 году. 56>Ханс Спеманн, который первым популяризировал термин «первичная нервная индукция» в отношении первой дифференцировки эктодермы в нервную ткань во время нейруляции. Это было названо «первичным», потому что считалось первым индукционным событием в эмбриогенезе. Эксперимент, получивший Нобелевскую премию, был проведен его ученицей Хильдой Мангольд. Эктодерма из области дорсальной губы бластопора развивающегося эмбриона саламандры была трансплантирована в другой эмбрион, и эта ткань «организатор» «индуцировала» образование полной вторичной оси, изменяя окружающую ткань в исходном эмбрионе с эктодермальной ткани на нервную ткань. Поэтому ткань донорского эмбриона называли индуктором, потому что она вызывала изменение. Важно отметить, что хотя организатором является дорсальная губа бластопора, это не один набор клеток, а скорее постоянно меняющаяся группа клеток, которые мигрируют по дорсальной губе бластопора, образуя апикально суженные бутылочные клетки. В любой момент времени во время гаструляции будут разные клетки, составляющие организатор.
Последующая работа ученых 20 века над индукторами продемонстрировала, что не только дорсальная губа бластопора может действовать как индуктор, но также может и огромное количество других, казалось бы, не связанных между собой предметов. Это началось, когда Йоханнес Хольтфретер обнаружил, что вареная эктодерма все еще может вызывать. Такие разнообразные предметы, как низкий pH, циклический AMP, даже пыль с пола, могут действовать как индукторы, вызывающие сильное беспокойство. Даже ткань, которая не могла образоваться при жизни, могла образоваться при кипячении. Другие предметы, такие как сало, воск, банановая кожура и свернувшаяся кровь лягушки, не вызывали. Охота за молекулой-индуктором на химической основе была занята молекулярными биологами, занимающимися вопросами развития, и обширная литература, посвященная объектам, обладающим способностью к индуктору, продолжала расти. Совсем недавно молекула-индуктор была приписана генам, а в 1995 году прозвучал призыв каталогизировать все гены, участвующие в первичной нейронной индукции, и все их взаимодействия, чтобы определить «молекулярную природу организатора Спемана». Некоторые другие белки и факторы роста также использовались в качестве индукторов, включая растворимые факторы роста, такие как морфогенетический белок кости, и потребность в «ингибирующих сигналах», таких как noggin и фоллистатин.
Еще до того, как термин индукция был популяризирован, несколько авторов, начиная с Ганса Дриша в 1894 году, предположили, что первичная нервная индукция может быть механической по своей природе. Основанная на механохимии модель первичной нейронной индукции была предложена в 1985 году Бродландом Гордоном. Было показано, что реальная физическая волна сокращения происходит из точного местоположения организатора Спемана, который затем пересекает предполагаемый нервный эпителий, и полная рабочая модель того, как первичная нейронная индукция была предложена в 2006 году. поле, чтобы рассмотреть возможность того, что первичная нейронная индукция может быть инициирована механическими воздействиями. Полное объяснение первичной нейронной индукции еще предстоит найти.
По мере того, как нейруляция продолжается после индукции, клетки нервной пластинки становятся высококолончатыми и могут быть идентифицированы при микроскопии как отличные от окружающей презумптивной эпителиальной эктодермы ( эпибластическая энтодерма у амниот). Клетки перемещаются в стороны и от центральной оси и принимают форму усеченной пирамиды. Эта форма пирамиды достигается за счет тубулина и актина в апикальной части клетки, которая сужается при движении. Вариация формы клеток частично определяется расположением ядра внутри клетки, вызывая выпуклость в областях клеток, что приводит к изменению высоты и формы клетки. Этот процесс известен как апикальное сужение. В результате дифференцирующаяся нервная пластинка уплощается, что особенно заметно у саламандр, когда ранее округлая гаструла превращается в округлый шар с плоской вершиной. См. Нервная пластинка
Процесс складывания плоской нервной пластинки в цилиндрическую нервную трубку называется первичной нейруляцией . В результате изменения клеточной формы нервная пластинка образует (MHP). Расширяющийся эпидермис оказывает давление на MHP и заставляет нервную пластинку складываться, что приводит к нервным складкам и созданию нервной борозды. Формируются нервные складки (DLHP), и давление на этот шарнир заставляет нервные складки встречаться и сливаться по средней линии. Слияние требует регуляции молекул клеточной адгезии. Нервная пластинка переключается с экспрессии E-кадгерина на экспрессию N-кадгерина и N-CAM, чтобы распознавать друг друга как одну и ту же ткань и закрывать трубку. Это изменение экспрессии останавливает связывание нервной трубки с эпидермисом. Складывание нервной пластинки - сложный этап.
Хорда играет важную роль в развитии нервной трубки. Перед нейруляцией, во время миграции клеток эпибластной энтодермы к гипобластической энтодерме, хордовый отросток открывается в дугу, называемую хордовой пластинкой, и прикрепляется к нейроэпителию нервной пластинки. Затем хордальная пластинка служит якорем для нервной пластинки и подталкивает два края пластинки вверх, удерживая при этом среднюю часть. Некоторые из нотходральных клеток включаются в центральную часть нервной пластинки, чтобы позже сформировать пластинку дна нервной трубки. Пластина хорды отделяется и образует твердую хорду.
Складывание нервной трубки в настоящую трубку не происходит сразу. Вместо этого он начинается примерно на уровне четвертого сомита на стадии Карнеги 9 (около 20-го дня эмбрионального развития у людей ). Боковые края нервной пластинки соприкасаются по средней линии и соединяются вместе. Это продолжается как краниально (к голове), так и каудально (к хвосту). Отверстия, которые образуются в краниальной и каудальной областях, называются краниальными и каудальными нейропорами . У эмбрионов человека краниальные нейропоры закрываются примерно на 24 день, а каудальные нейропоры - на 28 день. Неспособность закрытия краниальных (верхних) и каудальных (нижних) нейропор приводит к состояниям, называемым анэнцефалией и spina bifida соответственно. Кроме того, неспособность нервной трубки закрыться по всей длине тела приводит к состоянию, называемому рахишизис.
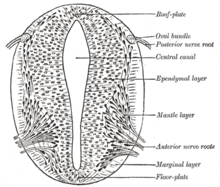 Поперечный срез нервной трубки, показывающий пластину пола и пластину крыши
Поперечный срез нервной трубки, показывающий пластину пола и пластину крыши Модель французского флага, где стадии развития управляются градиентами генных продуктов, несколько генов считаются важными для индукции паттернов в открытой нервной пластинке, особенно для развития нейрогенных плакод. Эти плакоды впервые становятся очевидными гистологически в открытой нервной пластинке. После того, как sonic hedgehog (SHH) передача сигналов от хорды индуцирует его образование, пластинка дна зарождающейся нервной трубки также секретирует SHH. После закрытия нервная трубка образует базальную или нижнюю пластину и крышу или крыловую пластину в ответ на комбинированное воздействие SHH и факторов, включая BMP4, секретируемые пластина крыши. Базальная пластинка образует большую часть вентральной части нервной системы, включая двигательную часть спинного мозга и ствол головного мозга; пластинка крыльев формирует дорсальные части, предназначенные в основном для сенсорной обработки.
Дорсальный эпидермис экспрессирует BMP4 и BMP7. Верхняя пластинка нервной трубки реагирует на эти сигналы экспрессией большего количества сигналов BMP4 и других сигналов трансформирующего фактора роста бета (TGF-β) с образованием дорсального / вентрального градиента между нервной трубкой. Нотохорд выражает SHH. Плита пола реагирует на SHH, создавая свою собственную SHH и образуя градиент. Эти градиенты допускают дифференциальную экспрессию факторов транскрипции.
Закрытие нервной трубки полностью не изучены. Закрытие нервной трубки зависит от вида. У млекопитающих закрытие происходит путем встречи в нескольких точках, которые затем закрываются вверх и вниз. У птиц закрытие нервной трубки начинается в одной точке среднего мозга и перемещается вперед и назад.
При вторичной нейруляции нервная эктодерма и некоторые клетки энтодермы образуют спинномозговой канатик. Медуллярный канатик уплотняется, отделяется и затем образует полости. Затем эти полости сливаются в одну трубку. Вторичная нейруляция наблюдается в заднем отделе у большинства животных, но у птиц она выражена лучше. Трубки первичной и вторичной нейруляции в конечном итоге соединяются примерно на шестой неделе развития.
Передняя часть нервной трубки образует три основные части головного мозга: передний мозг (передний мозг ), средний мозг (средний мозг ) и задний мозг (ромбовидный мозг ). Эти структуры первоначально появляются сразу после закрытия нервной трубки в виде выпуклостей, называемых мозговыми пузырьками, в паттерне, определяемом передне-задними генами формирования паттерна, включая гены Hox, другие факторы транскрипции такие как гены Emx, Otx и Pax, и секретируемые сигнальные факторы, такие как факторы роста фибробластов (FGFs) и Wnts. Эти мозговые пузырьки далее делятся на субрегионы. Передний мозг дает начало конечному мозгу и промежуточному мозгу, а ромбовидный мозг порождает передний мозг и продолговатый мозг. Задний мозг, который является эволюционно наиболее древней частью хордовых головного мозга, также делится на различные сегменты, называемые ромбомерами. Ромбомеры генерируют многие из наиболее важных нервных цепей, необходимых для жизни, включая те, которые контролируют дыхание и частоту сердечных сокращений, и производят большую часть черепных нервов. Клетки нервного гребня образуют ганглии над каждым ромбомером. Ранняя нервная трубка в основном состоит из зародышевого нейроэпителия, позже названного желудочковой зоной, который содержит первичные нервные стволовые клетки, называемые клетки радиальной глии и служит основным источником нейронов, образующихся в процессе развития мозга в процессе нейрогенеза.
Параксиальной мезодермы окружающие хорду по бокам разовьются в сомиты (будущие мышцы, кости и будут способствовать формированию конечностей позвоночных ).
ткань, называемая нервным гребнем, которая расположена на самых краях боковых пластинок складывающейся нервной трубки, отделяется от нервной трубки и мигрирует, чтобы стать множеством различных, но важных клеток.
Клетки нервного гребня будут мигрировать через эмбрион и давать начало нескольким клеточным популяциям, включая пигментные клетки и клетки периферических органов. нервная система.
Нарушение нейруляции, особенно нарушение закрытия нервной трубки, являются одними из наиболее распространенных и приводящих к инвалидности врожденных дефектов у людей, возникающих в примерно 1 из каждых 500 живорождений. Неспособность закрыться ростральным концом нервной трубки приводит к анэнцефалии или отсутствию развития мозга и чаще всего приводит к летальному исходу. Отсутствие закрытия хвостового конца нервной трубки вызывает состояние, известное как spina bifida, при котором спинной мозг не закрывается.