
Войти
Пути передачи сигналов Wnt : группа путей передачи сигнала, которые начинаются с белков, которые передают сигналы в клетку через рецепторы на поверхности клетки. Имя Wnt - это портмоне, созданное из имен Wingless и Int-1. Пути передачи сигналов Wnt используют либо связь между соседними клетками (паракрин ), либо связь между теми же клетками (аутокринные ). Они высоко эволюционно консервативны у животных, что означает, что они подобны для всех видов животных, от плодовых мух до людей.
Были охарактеризованы три сигнальных пути Wnt: канонический путь Wnt, путь неканонической планарной клеточной полярности и неканонический путь Wnt / кальция . Все три пути активируются связыванием Wnt-белка лиганда с рецептором семейства Frizzled , который передает биологический сигнал на Disheveled белок внутри клетки. Канонический путь Wnt приводит к регуляции транскрипции гена и, как полагают, частично негативно регулируется геном SPATS1. Неканонический путь плоской клеточной полярности регулирует цитоскелет, который отвечает за форму клетки. Неканонический путь Wnt / кальций регулирует кальций внутри клетки.
Передача сигналов Wnt была сначала идентифицирована по его роли в канцерогенезе, а затем по его функции в эмбриональном развитии. Эмбриональные процессы, которые он контролирует, включают формирование осей тела паттерна, определение клеточной судьбы, пролиферацию клеток и миграцию клеток. Эти процессы необходимы для правильного формирования важных тканей, включая кости, сердце и мышцы. Его роль в эмбриональном развитии была обнаружена, когда генетические мутации в белках пути Wnt приводили к образованию аномальных эмбрионов плодовой мухи . Передача сигналов Wnt также контролирует регенерацию ткани в костном мозге, коже и кишечнике взрослых. Более поздние исследования показали, что гены, ответственные за эти аномалии, также влияют на развитие рака груди у мышей.
Клиническое значение этого пути было продемонстрировано мутациями, которые приводят к различным заболеваниям, включая рак груди и рак простаты, глиобластому, сахарный диабет 2 типа и другие. В последние годы исследователи сообщили о первом успешном использовании ингибиторов пути Wnt на мышиных моделях заболевания.
На открытие передачи сигналов Wnt повлияли исследование онкогенных (вызывающих рак) ретровирусов. В 1982 году Роэль Нусс и Гарольд Вармус заразили мышей вирусом опухоли молочной железы мышей, чтобы мутировать гены мыши, чтобы увидеть, какие мутированные гены могут вызывать опухоли груди. Они идентифицировали новый протоонкоген мыши, который они назвали int1 (интеграция 1).
Int1 является высококонсервативным у многих видов, включая человека и Drosophila. Его присутствие в Д. melanogaster привел исследователей к открытию в 1987 году, что ген int1 у Drosophila на самом деле был уже известным и охарактеризованным геном Drosophila, известным как Wingless (Wg). Поскольку предыдущее исследование Кристиан Нюсслейн-Фольхард и Эрика Вишауса (которое принесло им Нобелевскую премию по физиологии и медицине в 1995 году) уже установило функцию Wg. Поскольку ген полярности сегмента участвует в формировании оси тела во время эмбрионального развития, исследователи определили, что int1 млекопитающих, обнаруженный у мышей, также участвует в эмбриональном развитии.
Продолжение исследований привело к открытию дополнительных генов, связанных с int1; однако, поскольку эти гены не были идентифицированы таким же образом, как int1, номенклатура гена int была неадекватной. Таким образом, семейство int / Wingless стало семейством Wnt, а int1 стало Wnt1. Название Wnt представляет собой портманто из int и Wg и означает «сайт интеграции, связанный с бескрылыми».
 Структура кристаллического белка Wnt8 и богатый цистеином домен Frizzled 8
Структура кристаллического белка Wnt8 и богатый цистеином домен Frizzled 8 Wnt включает разнообразное семейство секретируемых липид -модифицированных сигнальных гликопротеинов, которые имеют длину 350-400 аминокислот. Модификация липидов всех Wnts представляет собой пальмитолеоилирование одного полностью консервативного остатка серина. Пальмитолеоилирование необходимо, потому что это требуется для Wnt для связывания со своим белком-носителем Wntless (WLS), чтобы он мог транспортироваться к плазматической мембране для секреции, а также позволяет белку Wnt связываться со своим рецептором. Frizzled Wnt белки. подвергаются гликозилированию, которое присоединяет углевод для обеспечения надлежащей секреции. В передаче сигналов Wnt эти белки действуют как лиганды для активации различных путей Wnt паракринным и аутокринным путями.
Эти белки высоко консервативны у разных видов. Их можно найти у мышей, людей, Xenopus, рыбок данио, Drosophila и многих других.
| Виды | Белки Wnt | ||
|---|---|---|---|
| Homo sapiens | WNT1, WNT2, WNT2B, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16 | ||
| Mus musculus (идентичные белки как у H. sapiens) | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt9A, Wnt9B, Wnt10Ant | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt11R | |
| Wnt2B, Wnt2 rerio | Данио rerio | Данио rerio | , Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Drosophila | Wg, DWnt2, DWnt3 / 5, DW6nt 4, DWnt3 / 5, DW6nt 4, DWnt DWnt8, DWnt10 | ||
| Hydra | hyw nt1, hywnt5a, hywnt8, hywnt7, hywnt9 / 10a, hywnt9 / 10b, hywnt9 / 10c, hywnt11, hywnt16 | ||
| C. elegans | mom-2, lin-44, egl-20, cwn-1, cwn-2 |
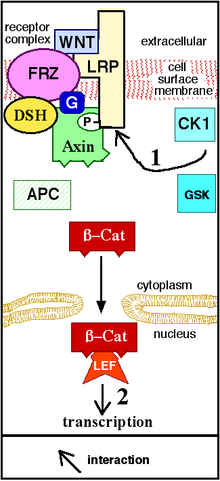 Рис. 2. Wnt связывается (активирует) рецептор. Аксин удален из «комплекса разрушения». β-Cat перемещается в ядро, связывается с фактором транскрипции на ДНК и активирует транскрипцию белка. «P» представляет фосфат.
Рис. 2. Wnt связывается (активирует) рецептор. Аксин удален из «комплекса разрушения». β-Cat перемещается в ядро, связывается с фактором транскрипции на ДНК и активирует транскрипцию белка. «P» представляет фосфат. . Фиг.1. Wnt не связывается с рецептором. Аксин, GSK и APC образуют «деструктивный комплекс», и β-Cat разрушается.
. Фиг.1. Wnt не связывается с рецептором. Аксин, GSK и APC образуют «деструктивный комплекс», и β-Cat разрушается. .
.
.
.
.
.
.
.
.
.
.
Передача сигнала Wnt начинается, когда белок Wnt связывается с N-концевым внеклеточным богатым цистеином доменом Рецептор семейства Frizzled (Fz). Эти рецепторы охватывают плазматическую мембрану семь раз и составляют отдельное семейство рецепторов, связанных с G-белком (GPCR). Однако для облегчения передачи сигналов Wnt могут потребоваться корецепторы наряду с взаимодействием между белком Wnt и рецептором Fz. Примеры включают белок, связанный с рецептором липопротеина (LRP ) -5/6, рецепторную тирозинкиназу (RTK) и ROR2. После активации рецептора сигнал посылается в фосфопротеин Disheveled (Dsh), который находится в цитоплазме. Этот сигнал передается через прямое взаимодействие между Fz и Dsh. Белки Dsh присутствуют во всех организмах, и все они имеют следующие высококонсервативные белковые домены : аминоконцевой домен DIX, центральный домен PDZ и карбоксиконцевой Домен DEP. Эти разные домены важны, потому что после Dsh сигнал Wnt может разветвляться на несколько путей, и каждый путь взаимодействует с другой комбинацией трех доменов.
Три лучших Охарактеризованные пути передачи сигналов Wnt представляют собой канонический путь Wnt, путь неканонической планарной клеточной полярности и неканонический путь Wnt / кальция. Как следует из их названия, эти пути относятся к одной из двух категорий: канонической или неканонической. Разница между категориями заключается в том, что канонический путь включает белок β-катенин, тогда как неканонический путь действует независимо от него.
 Канонический путь Wnt
Канонический путь Wnt Канонический путь Путь Wnt (или путь Wnt / β-катенина) - это путь Wnt, который вызывает накопление β-катенина в цитоплазме и его возможную транслокацию в ядро , чтобы действовать как транскрипционный коактиватор из факторов транскрипции, принадлежащих к семейству TCF / LEF. Без Wnt β-catenin не будет накапливаться в цитоплазме, поскольку комплекс деструкции обычно разрушает его. Этот комплекс деструкции включает следующие белки: Axin, аденоматозный полипоз coli (APC), протеинфосфатаза 2A (PP2A), киназа гликогенсинтазы 3 (GSK3) и казеинкиназа 1 α (CK1α). Он расщепляет β-катенин, направляя его на убиквитинирование, которое впоследствии отправляет его в протеасому для переваривания. Однако, как только Wnt связывает Fz и LRP5 /6, функция комплекса разрушения нарушается. Это связано с тем, что Wnt вызывает перемещение негативного регулятора Wnt, Axin, и деструктивного комплекса к плазматической мембране. Фосфорилирование другими белками в комплексе деструкции впоследствии связывает Axin с цитоплазматическим хвостом LRP5 / 6. Аксин де-фосфорилируется, его стабильность и уровни снижаются. Затем Dsh активируется посредством фосфорилирования, и его домены DIX и PDZ ингибируют активность GSK3 комплекса деструкции. Это позволяет β-катенину накапливаться и локализоваться в ядре, а затем вызывать клеточный ответ посредством трансдукции гена вместе с факторами транскрипции TCF / LEF (T-клеточный фактор / лимфоидный фактор). β-катенин задействует другие факторы. коактиваторы транскрипции, такие как BCL9, Pygopus и Parafibromin / Hyrax. Сложность транскрипционного комплекса, собираемого β-катенином, начинает проявляться благодаря новым высокопроизводительным исследованиям протеомики. Экстенсивность белков, взаимодействующих с β-катенином, усложняет наше понимание: β-катенин может непосредственно фосфорилироваться по Ser552 с помощью Akt, что вызывает его диссоциацию от межклеточных контактов и накопление в цитозоле, после чего 14-3- 3ζ взаимодействует с β-катенином (pSer552) и усиливает его ядерную транслокацию. Сообщалось, что BCL9 и Pygopus действительно обладают несколькими β-катенином - независимые функции (следовательно, вероятно, независимые от передачи сигналов Wnt).
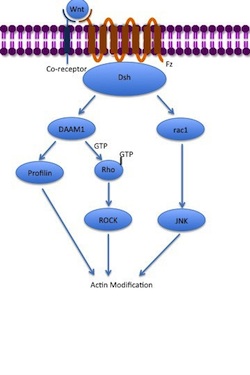 Неканонический путь PCP
Неканонический путь PCP Путь неканонической планарной полярности клеток (PCP) не включает β-катенин. Он не использует LRP-5/6 в качестве своего корецептора и, как предполагается, использует PTK7 или ROR2. Путь PCP активируется посредством связывания Wnt с Fz и его корецептором. Затем рецептор привлекает Dsh, который использует свои PDZ- и DIX-домены для образования комплекса с Disheveled-ассоциированным активатором морфогенеза 1 (DAAM1 ). Затем Daam1 активирует небольшой G-белок Rho посредством фактора обмена гуанина. Rho активирует Rho-ассоциированную киназу (ROCK), которая является одним из основных регуляторов цитоскелета. Dsh также образует комплекс с rac1 и опосредует связывание профилина с актином. Rac1 активирует JNK и может также привести к полимеризации актина . Связывание профилина с актином может приводить к реструктуризации цитоскелета и гаструляции.
 Неканонический путь Wnt / кальция
Неканонический путь Wnt / кальция Неканонический путь Wnt / кальция также не включает β-катенин. Его роль состоит в том, чтобы помочь регулировать высвобождение кальция из эндоплазматического ретикулума (ER), чтобы контролировать уровни внутриклеточного кальция. Подобно другим путям Wnt, при связывании лиганда активированный рецептор Fz напрямую взаимодействует с Dsh и активирует специфические домены Dsh-белка. Домены, участвующие в передаче сигналов Wnt / кальция, представляют собой домены PDZ и DEP. Однако, в отличие от других путей Wnt, рецептор Fz напрямую взаимодействует с тримерным G-белком. Такая совместная стимуляция Dsh и G-белка может привести к активации либо PLC, либо cGMP-специфической PDE. Если PLC активирован, компонент плазматической мембраны PIP2 расщепляется на DAG и IP3. Когда IP3 связывает свой рецептор на ER, высвобождается кальций. Повышенные концентрации кальция и DAG могут активировать от Cdc42 до PKC. Cdc42 является важным регулятором формирования вентрального паттерна. Повышенный уровень кальция также активирует кальциневрин и CaMKII. CaMKII индуцирует активацию фактора транскрипции NFAT, который регулирует клеточную адгезию, миграцию и разделение тканей. Кальциневрин активирует киназу TAK1 и NLK, которые могут мешать передаче сигналов TCF / ß-катенин в каноническом пути Wnt. Однако, если PDE активируется, высвобождение кальция из ER ингибируется. PDE опосредует это посредством ингибирования PKG, что впоследствии вызывает ингибирование высвобождения кальция.
Бинарное различие канонических и неканонических сигнальных путей Wnt подверглось тщательному изучению и был предложен интегрированный, конвергентный путь Wnt. Некоторые доказательства этого были обнаружены для одного лиганда Wnt (Wnt5A). Доказательства конвергентного пути передачи сигналов Wnt, который демонстрирует интегрированную активацию передачи сигналов Wnt / Ca2 + и Wnt / ß-катенин для множественных лигандов Wnt, были описаны в линиях клеток млекопитающих.
Wnt передача сигналов также регулирует ряд других путей передачи сигналов, которые не были столь подробно изучены. Один из таких путей включает взаимодействие между Wnt и GSK3. Во время роста клеток Wnt может ингибировать GSK3, чтобы активировать mTOR в отсутствие β-катенина. Однако Wnt также может служить негативным регулятором mTOR посредством активации опухолевого супрессора TSC2, который активируется посредством взаимодействия Dsh и GSK3. Во время миогенеза Wnt использует PA и CREB для активации генов MyoD и Myf5. Wnt также действует вместе с Ryk и Src, позволяя регулировать отталкивание нейронов во время наведения аксонов. Wnt регулирует гаструляцию, когда CK1 служит ингибитором Rap1-ATPase, чтобы модулировать цитоскелет во время гаструляции. Дальнейшая регуляция гаструляции достигается, когда Wnt использует ROR2 вместе с путями CDC42 и JNK для регуляции экспрессии PAPC. Dsh также может взаимодействовать с aPKC, а также для того, чтобы контролировать полярность клеток и развитие цитоскелета микротрубочек. Хотя эти пути перекрываются с компонентами, связанными с передачей сигналов PCP и Wnt / Calcium, они считаются отдельными путями, потому что они вызывают разные ответы.
Для обеспечения правильного функционирования передача сигналов Wnt постоянно регулируется в нескольких точках на его сигнальных путях. Например, белки Wnt пальмитоилированы. Белок дикобраз опосредует этот процесс, что означает, что он помогает регулировать, когда лиганд Wnt секретируется, определяя, когда он полностью сформирован. Секреция дополнительно контролируется с помощью белков, таких как GPR177 (без wntless), и комплексов, таких как комплекс ретромера.
При секреции лиганд можно предотвратить достижение его рецептора за счет связывания белков, таких как стабилизаторы Dally и глипикан 3 (GPC3), которые ингибируют диффузию. В раковых клетках и гепарансульфатные цепи, и основной белок GPC3 участвуют в регуляции связывания Wnt и активации пролиферации клеток. Wnt распознает структуру гепарансульфата на GPC3, которая содержит IdoA2S и GlcNS6S, а 3-O-сульфатирование в GlcNS6S3S усиливает связывание Wnt с гепарансульфат-глипиканом. Богатый цистеином домен в N-доле GPC3 был идентифицирован как формирующий Wnt-связывающую гидрофобную бороздку, включающую фенилаланин-41, которая взаимодействует с Wnt. Блокирование связывающего домена Wnt с помощью нанотела, называемого HN3, может ингибировать активацию Wnt.
На рецепторе Fz связывание белков, отличных от Wnt, может противодействовать передаче сигналов. Конкретные антагонисты включают Dickkopf (Dkk), фактор ингибирования Wnt 1 (WIF-1), секретируемые Frizzled-связанные белки (SFRP), Цербер, Фрзб, Мудрый, СОСТ и Голая кутикула. Они представляют собой ингибиторы передачи сигналов Wnt. Однако другие молекулы также действуют как активаторы. Норрин и R-Spondin2 активируют передачу сигнала Wnt в отсутствие лиганда Wnt.
Взаимодействия между сигнальными путями Wnt также регулируют передачу сигналов Wnt. Как упоминалось ранее, путь Wnt / кальций может ингибировать TCF / β-катенин, предотвращая передачу сигналов канонического пути Wnt. Простагландин E2 является важным активатором канонического пути передачи сигналов Wnt. Взаимодействие PGE2 с его рецепторами E2 / E4 стабилизирует β-катенин посредством фосфорилирования, опосредованного цАМФ / PKA. Синтез PGE2 необходим для процессов, опосредованных передачей сигналов Wnt, таких как регенерация тканей и контроль популяции стволовых клеток у рыбок данио и мышей. Интересно, что неструктурированные области нескольких слишком больших белков с внутренним нарушением играют решающую роль в регуляции передачи сигналов Wnt.
Передача сигналов Wnt играет решающую роль в эмбриональном развитии. Он действует как у позвоночных, так и беспозвоночных, включая людей, лягушек, рыбок данио, C. elegans, Drosophila и другие. Впервые он был обнаружен в сегменте полярности Drosophila, где он помогает установить переднюю и заднюю полярности. Он вовлечен в другие процессы развития. Как предполагает его функция у Drosophila, он играет ключевую роль в формировании оси тела, особенно в формировании переднезадней и дорсовентральной осей. Он участвует в индукции дифференцировки клеток, чтобы стимулировать образование важных органов, таких как легкие и яичники. Wnt дополнительно обеспечивает развитие этих тканей посредством правильной регуляции пролиферации клеток и миграции. Функции передачи сигналов Wnt можно разделить на формирование осевого паттерна, спецификацию клеточной судьбы, пролиферацию и миграцию клеток.
На раннем этапе развития эмбриона формирование основных осей тела является решающим этапом в установлении общего плана организма. Оси включают переднезаднюю ось, дорсовентральную ось и правую-левую ось. Передача сигналов Wnt участвует в формировании переднезадней и дорсовентральной (DV) осей. Активность передачи сигналов Wnt в передне-заднем развитии можно наблюдать у млекопитающих, рыб и лягушек. У млекопитающих примитивная полоса и другие окружающие ткани продуцируют морфогенные соединения Wnts, BMP, FGFs, Nodal и ретиноид. acid для образования задней области во время поздней гаструлы. Эти белки образуют градиенты концентрации. Области наибольшей концентрации определяют заднюю область, а области наименьшей концентрации указывают переднюю область. У рыб и лягушек β-catenin, продуцируемый канонической передачей сигналов Wnt, вызывает образование центров организации, которые, наряду с BMPs, вызывают образование задних отделов. Участие Wnt в формировании оси DV можно увидеть в активности образования Spemann-организатора, который устанавливает дорсальную область. Каноническая передача сигналов Wnt продукция β-catenin индуцирует образование этого организатора через активацию генов twin и siamois. Точно так же при гаструляции птиц клетки серпа Коллера экспрессируют различные гены-маркеры мезодермы, которые позволяют дифференцированное движение клеток во время образования примитивной полоски. Передача сигналов Wnt, активируемая FGFs, ответственна за это движение.
Передача сигналов Wnt также участвует в формировании осей специфических частей тела и систем органов на более позднем этапе развития. У позвоночных sonic hedgehog (Shh) и градиенты морфогенетических сигналов Wnt устанавливают дорсовентральную ось центральной нервной системы во время формирования осевого паттерна нервной трубки. Высокий уровень передачи сигналов Wnt устанавливает дорсальную область, тогда как высокий уровень передачи сигналов Shh указывает вентральную область. Wnt участвует в формировании DV центральной нервной системы посредством его участия в ведении аксонов. Белки Wnt направляют аксоны спинного мозга в передне-заднем направлении. Wnt также участвует в формировании оси DV конечности. В частности, Wnt7a помогает создавать дорсальный паттерн развивающейся конечности.
В модели развития волн эмбриональной дифференцировки Wnt играет критическую роль как часть сигнального комплекса в компетентных клетках, готовых к дифференцировке. Wnt реагирует на активность цитоскелета, стабилизируя начальные изменения, создаваемые проходящей волной сжатия или расширения, и одновременно сигнализирует ядру с помощью различных сигнальных путей о том, в какой волне участвовала отдельная клетка. Таким образом, активность Wnt усиливается. механическая передача сигналов, которая происходит во время развития.
Спецификация клеточной судьбы или дифференцировка клеток - это процесс, при котором недифференцированные клетки могут стать более специализированным типом клеток. Передача сигналов Wnt индуцирует дифференцировку плюрипотентных стволовых клеток в мезодерму и эндодерму клетки-предшественники. Эти клетки-предшественники далее дифференцируются в типы клеток, такие как эндотелиальные, сердечные и сосудистые гладкие мышцы. Передача сигналов Wnt индуцирует образование крови из стволовых клеток. В частности, Wnt3 приводит к коммитированным клеткам мезодермы с гематопоэтическим потенциалом. Wnt1 противодействует нейральной дифференцировке и является основным фактором самообновления нервных стволовых клеток. Это позволяет регенерацию клеток нервной системы, что является дополнительным свидетельством их роли в стимулировании пролиферации нервных стволовых клеток. Передача сигналов Wnt участвует в определении зародышевых клеток, спецификации ткани кишечника, развитии волосяного фолликула, развитии легочной ткани, дифференцировке стволовых клеток нервного гребня, развитие нефрона, развитие яичников и определение пола. Передача сигналов Wnt также препятствует формированию сердца, и было показано, что ингибирование Wnt является критическим индуктором сердечной ткани во время развития, и низкомолекулярные ингибиторы Wnt обычно используются для производства кардиомиоцитов из плюрипотентных стволовых клеток.
Для массовой дифференцировки клеток, необходимой для образования указанных клеточных тканей различных организмов, должны иметь место пролиферация и рост эмбриональных стволовых клеток. Этот процесс опосредуется канонической передачей сигналов Wnt, которая увеличивает ядерный и цитоплазматический β-catenin. Повышенный уровень β-катенина может инициировать активацию транскрипции белков, таких как циклин D1 и c-myc, которые контролируют фазу от G1 до S переход в клеточном цикле. Вступление в S-фазу вызывает репликацию ДНК и в конечном итоге митоз, которые ответственны за пролиферацию клеток. Это увеличение пролиферации напрямую связано с дифференцировкой клеток, потому что по мере пролиферации стволовых клеток они также дифференцируются. Это обеспечивает общий рост и развитие определенных тканевых систем во время эмбрионального развития. Это очевидно в таких системах, как система кровообращения, где Wnt3a приводит к пролиферации и экспансии гемопоэтических стволовых клеток, необходимых для образования красных кровяных телец.
Биохимия раковых стволовых клеток немного отличается от что других опухолевых клеток. Эти так называемые Wnt-зависимые клетки захватывают и зависят от постоянной стимуляции пути Wnt, чтобы способствовать их неконтролируемому росту, выживанию и миграции. При раке передача сигналов Wnt может стать независимой от регулярных стимулов посредством мутаций в последующих онкогенах и генах-супрессорах опухолей, которые становятся постоянно активированными, даже если нормальный рецептор не получил сигнала. β-катенин связывается с факторами транскрипции, такими как белок TCF4, и в сочетании молекулы активируют необходимые гены. LF3 сильно ингибирует это связывание in vitro в клеточных линиях и снижает рост опухоли на моделях мышей. Это препятствовало репликации и уменьшало их способность к миграции, не затрагивая здоровые клетки. После лечения не осталось раковых стволовых клеток. Это открытие явилось результатом «рационального дизайна лекарств » с использованием технологий AlphaScreens и ELISA.
Миграция клеток во время эмбрионального развития позволяет устанавливать оси тела, формирование тканей, индукцию конечностей и ряд других процессов. Передача сигналов Wnt помогает опосредовать этот процесс, особенно во время конвергентного расширения. Передача сигналов как от пути Wnt PCP, так и от канонического пути Wnt необходима для правильного конвергентного расширения во время гаструляции. Конвергентное удлинение дополнительно регулируется путем Wnt / Calc, который блокирует конвергентное удлинение при активации. Передача сигналов Wnt также индуцирует миграцию клеток на более поздних стадиях развития посредством контроля поведения миграции нейробластов, клеток нервного гребня, миоцитов и клеток трахеи. 292>
Передача сигналов Wnt участвует в другом ключевом процессе миграции, известном как эпителиально-мезенхимальный переход (EMT). Этот процесс позволяет эпителиальным клеткам превращаться в мезенхимальные клетки, так что они больше не удерживаются на месте на ламинине. Он включает подавление регуляции кадгерина, так что клетки могут отделяться от ламинина и мигрировать. Передача сигналов Wnt является индуктором ЭМП, особенно при развитии молочных желез.
 Диаграмма, иллюстрирующая взаимодействие между путями передачи сигналов Wnt и инсулина
Диаграмма, иллюстрирующая взаимодействие между путями передачи сигналов Wnt и инсулина Инсулин является пептидным гормоном участвует в глюкозе гомеостазе в определенных организмах. В частности, это приводит к положительной регуляции переносчиков глюкозы в клеточной мембране, чтобы увеличить захват глюкозы из кровотока. Этот процесс частично опосредуется активацией передачи сигналов Wnt / β-катенина, что может повысить чувствительность клетки к инсулину. В частности, Wnt10b является белком Wnt, который увеличивает эту чувствительность в клетках скелетных мышц.
С момента своего первоначального открытия передача сигналов Wnt была связана с рак. Когда был открыт Wnt1, он был впервые идентифицирован как прото- онкоген в мышиной модели рака груди. Тот факт, что Wnt1 является гомологом Wg, показывает, что он участвует в эмбриональном развитии, которое часто требует быстрого деления и миграции клеток. Неправильная регуляция этих процессов может привести к развитию опухоли за счет избыточной пролиферации клеток.
Каноническая активность пути Wnt участвует в развитии доброкачественных и злокачественных опухолей молочной железы. Его присутствие выявляется по повышенным уровням β-катенина в ядре и / или цитоплазме, что может быть обнаружено с помощью иммуногистохимического окрашивания и вестерн-блоттинга. Повышенная экспрессия β-катенина коррелирует с плохим прогнозом у пациентов с раком груди. Это накопление может быть связано с такими факторами, как мутации в β-катенине, дефицит комплекса разрушения β-катенина, чаще всего из-за мутаций в структурно неупорядоченных областях APC, сверхэкспрессии лигандов Wnt, потеря ингибиторов и т. Д. / или снижение активности регуляторных путей (таких как путь Wnt / кальций). Опухоли груди могут метастазировать из-за вовлечения Wnt в EMT. Исследования, посвященные метастазам базальноподобного рака молочной железы в легкие, показали, что подавление передачи сигналов Wnt / β-катенина может предотвращать ЕМТ, что может ингибировать метастазирование.
Передача сигналов Wnt участвует в развитии других видов рака. Изменения экспрессии CTNNB1, который является геном, кодирующим β-катенин, можно измерить в груди, колоректальном, меланоме, простате, легкого и другие виды рака. Повышенная экспрессия белков-лигандов Wnt, таких как Wnt1, Wnt2 и Wnt7A, наблюдалась при развитии глиобластомы, рака пищевода и рака яичников соответственно. Другие белки, которые вызывают несколько типов рака при отсутствии надлежащего функционирования, включают ROR1, ROR2, SFRP4, Wnt5A, WIF1 и белки семейства TCF / LEF.
Связь между PGE2 и Wnt предполагает, что связанное с хроническим воспалением повышение уровня PGE2 может приводить к активации пути Wnt в различных тканях, что приводит к канцерогенезу.
Сахарный диабет 2 типа является распространенным заболеванием, вызывающим снижение секреции инсулина и повышение инсулинорезистентности на периферии. Это приводит к повышению уровня глюкозы в крови или гипергликемии, которая может быть фатальной при отсутствии лечения. Поскольку передача сигналов Wnt участвует в чувствительности к инсулину, это может быть связано с нарушением его пути. Сверхэкспрессия Wnt5b, например, может увеличивать восприимчивость из-за его роли в адипогенезе, поскольку ожирение и диабет II типа имеют высокую коморбидность. Передача сигналов Wnt является сильным активатором митохондриального биогенеза. Это приводит к увеличению производства активных форм кислорода (ROS), которые, как известно, вызывают повреждение ДНК и клеток. This ROS-induced damage is significant because it can cause acute hepatic insulin resistance, or injury-induced insulin resistance. Mutations in Wnt signaling-associated transcription factors, such as TCF7L2, are linked to increased susceptibility.