
Войти
| Различать до и после синапса |
|---|
 ↓ К постсинаптическому нейрону ↓ Из пресинаптического нейрона Транспортер нейротрансмиттера Рецептор нейротрансмиттера Передача нейротрансмиттера Синаптическая щель { ↓ К постсинаптическому нейрону ↓ Из пресинаптического нейрона Транспортер нейротрансмиттера Рецептор нейротрансмиттера Передача нейротрансмиттера Синаптическая щель { |
| «Соединение, связывающее нейрон с нейроном, - это синапс. Сигнал течет в одном направлении, от пресинаптического нейрона к постсинаптическому нейрону через синапс, который действует как переменный аттенюатор». Вкратце, направление потока сигнала определяет префикс задействованных синапсов. |
В нервной системе, A синапс представляет собой структуру, которая позволяет нейрон (или нервную клетку), чтобы передать электрический или химический сигнал другого нейрону или к целевым эффекторным клеткам.
Синапсы необходимы для передачи нервных импульсов от одного нейрона к другому. Нейроны специализируются на передаче сигналов отдельным клеткам-мишеням, и синапсы являются средством, с помощью которого они это делают. В синапсе плазматическая мембрана нейрона, передающего сигнал ( пресинаптического нейрона), вступает в тесное соприкосновение с мембраной целевой ( постсинаптической) клетки. И пресинаптические, и постсинаптические сайты содержат обширные массивы молекулярных механизмов, которые связывают две мембраны вместе и осуществляют процесс передачи сигналов. Во многих синапсах пресинаптическая часть расположена на аксоне, а постсинаптическая часть - на дендрите или соме. Астроциты также обмениваются информацией с синаптическими нейронами, отвечая на синаптическую активность и, в свою очередь, регулируя нейротрансмиссию. Синапсы (по крайней мере, химические синапсы) стабилизируются по положению за счет молекул синаптической адгезии (SAM), выступающих как из пре-, так и из постсинаптического нейрона и слипающихся вместе, где они перекрываются; SAM также могут способствовать образованию и функционированию синапсов.
Некоторые авторы обобщают концепцию синапса, чтобы включить связь нейрона с любым другим типом клеток, например с моторной клеткой, хотя такие ненейрональные контакты могут называться соединениями (исторически более старый термин). Знаменательное исследование Сэнфорда Палея продемонстрировало существование синапсов.
Сантьяго Рамон-и-Кахаль предположил, что нейроны не непрерывны по всему телу, но все же взаимодействуют друг с другом, идея, известная как доктрина нейронов. Слово «синапс» было введено в 1897 году английским нейрофизиологом Чарльзом Шеррингтоном в « Учебнике физиологии» Майкла Фостера. Шеррингтон изо всех сил пытался найти хороший термин, который подчеркивал бы союз между двумя отдельными элементами, а сам термин «синапс» был предложен английским классиком Артуром Вуллгаром Верраллом, другом Фостера. Слово произошло от греческого синапсиса ( συνάψις), что означает «соединение», которое, в свою очередь, происходит от συνάπτεὶν ( συν («вместе») и ἅπτειν («закреплять»)).
Однако, хотя синаптическая щель оставалась теоретической конструкцией и иногда описывалась как разрыв между смежными окончаниями аксонов и дендритами или клеточными телами, гистологические методы с использованием лучших световых микроскопов не могли визуально разрешить их разделение, которое, как теперь известно, примерно 20нм. Электронный микроскоп понадобился в 1950-х годах, чтобы показать более тонкую структуру синапса с его отдельными параллельными пре- и постсинаптическими мембранами и отростками, а также щель между ними.
 Пример химического синапса путем высвобождения нейромедиаторов, таких как ацетилхолин или глутаминовая кислота.
Пример химического синапса путем высвобождения нейромедиаторов, таких как ацетилхолин или глутаминовая кислота. Есть два принципиально разных типа синапсов:
Синаптическая связь отличается от эпаптической связи, при которой связь между нейронами происходит через непрямые электрические поля.
Autapse представляет собой химический или электрический синапс, который образуется, когда аксон одного нейрона синапсы на дендритах одного и того же нейрона.
Синапсы можно классифицировать по типу клеточных структур, служащих пре- и постсинаптическими компонентами. Подавляющее большинство синапсов в нервной системе млекопитающих представляют собой классические аксо-дендритные синапсы (синапсы аксонов на дендрите), однако существует множество других механизмов. Они включают, но не ограничиваются аксо-аксонального, дендропарк-дендритные, аксо-секреторной, сомато-дендритные, дендропарк-соматические и сомато-соматические синапсы.
Аксон может синапсировать с дендритом, с телом клетки или с другим аксоном или концом аксона, а также в кровоток или диффузно в соседнюю нервную ткань.
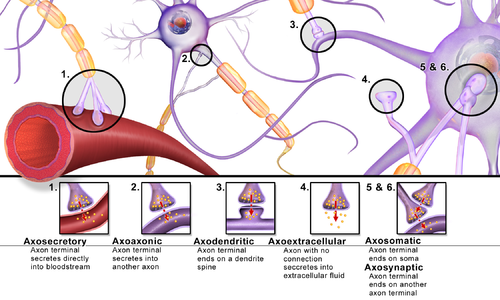 Различные типы синапсов
Различные типы синапсов Широко признано, что синапсы играют роль в формировании памяти. Поскольку нейротрансмиттеры активируют рецепторы в синаптической щели, связь между двумя нейронами усиливается, когда оба нейрона активны одновременно, в результате сигнальных механизмов рецептора. Считается, что сила двух связанных нейронных путей приводит к хранению информации, что приводит к памяти. Этот процесс усиления синапсов известен как долговременная потенциация.
Изменяя высвобождение нейротрансмиттеров, можно контролировать пластичность синапсов в пресинаптической клетке. Постсинаптическая клетка может регулироваться путем изменения функции и количества ее рецепторов. Изменения в постсинаптической передаче сигналов чаще всего связаны с зависимым от рецептора N-метил-d-аспарагиновой кислоты (NMDAR) долгосрочным потенцированием (LTP) и долгосрочным подавлением (LTD) из-за притока кальция в постсинаптический клетки, которые являются наиболее анализируемыми формами пластичности возбуждающих синапсов.
По техническим причинам синаптическая структура и функция исторически изучались на необычно больших модельных синапсах, например:
Функция нейронов зависит от полярности клеток. Отличительная структура нервных клеток позволяет потенциалам действия перемещаться направленно (от дендритов к телу клетки вниз по аксону), и эти сигналы затем принимаются и переносятся постсинаптическими нейронами или эффекторными клетками. Нервные клетки давно используются в качестве моделей клеточной поляризации, и особый интерес представляют механизмы, лежащие в основе поляризованной локализации синаптических молекул. Передача сигналов PIP2, регулируемая IMPase, играет важную роль в синаптической полярности.
Фосфоинозитиды ( PIP, PIP2 и PIP3 ) - это молекулы, которые, как было показано, влияют на полярность нейронов. Ген ( ttx-7) был идентифицирован у Caenorhabditis elegans, который кодирует мио- инозитолмонофосфатазу (IMPase), фермент, который продуцирует инозит путем дефосфорилирования инозитолфосфата. Организмы с мутантными генами ttx-7 демонстрировали дефекты поведения и локализации, которые были устранены экспрессией IMPase. Это привело к выводу, что ИМФаза необходима для правильной локализации синаптических белковых компонентов. Ген egl-8 кодирует гомолог фосфолипазы C β (PLCβ), фермента, расщепляющего PIP2. Когда мутанты ttx -7 также имели мутантный ген egl-8, дефекты, вызванные дефектным геном ttx-7, были в значительной степени обращены. Эти результаты предполагают, что передача сигналов PIP2 устанавливает поляризованную локализацию синаптических компонентов в живых нейронах.
Модуляция высвобождения нейротрансмиттеров рецепторами, связанными с G-белками (GPCR), является важным пресинаптическим механизмом регуляции синаптической передачи. Активация GPCR, расположенных на пресинаптическом конце, может снизить вероятность высвобождения нейромедиатора. Эта пресинаптическая депрессия включает активацию G-белков Gi / o- типа, которые опосредуют различные механизмы ингибирования, включая ингибирование потенциал-управляемых кальциевых каналов, активацию калиевых каналов и прямое ингибирование процесса слияния везикул. Эндоканнабиноиды, синтезируемые в постсинаптических нейронных элементах и высвобождаемые из них, и их родственные рецепторы, включая рецептор (GPCR) CB1, расположенный на пресинаптическом конце, участвуют в этой модуляции ретроградным сигнальным процессом, в котором эти соединения синтезируются и высвобождаются. от постсинаптических нейронных элементов и возвращаются к пресинаптическому окончанию, чтобы воздействовать на рецептор CB1 для краткосрочной (STD) или долгосрочной синаптической депрессии (LTD), которая вызывает кратковременное или длительное снижение высвобождения нейромедиатора.

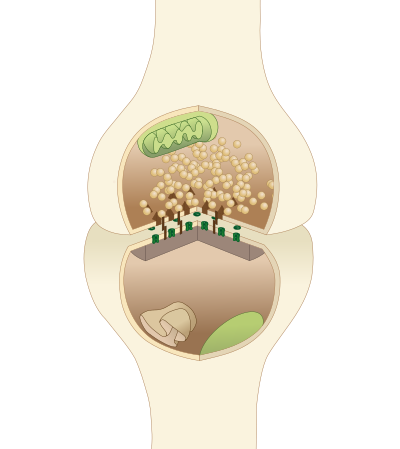
Типичный синапс центральной нервной системы

Синапс и цикл синаптических пузырьков

Основные элементы в химической синаптической передаче
