
Войти
Полярность ячейки относится к пространственным различиям в форме, структуре и функциях внутри ячейки . Почти все типы клеток обладают той или иной формой полярности, которая позволяет им выполнять специализированные функции. Классические примеры поляризованных клеток описаны ниже, включая эпителиальные клетки с апикально-базальной полярностью, нейроны, в которых сигналы распространяются в одном направлении от дендритов к аксонам и мигрирующие ячейки. Кроме того, полярность клеток важна во время многих типов асимметричного деления клеток для установления функциональной асимметрии между дочерними клетками.
Многие ключевые молекулярные игроки, участвующие в полярности клеток, хорошо законсервированы. Например, в клетках многоклеточных животных комплекс PAR-3 / PAR-6 / aPKC играет фундаментальную роль в полярности клеток. Хотя биохимические характеристики могут различаться, некоторые из основных принципов, такие как отрицательная и / или положительная обратная связь между различными молекулами, являются общими и важными для многих известных систем полярности.
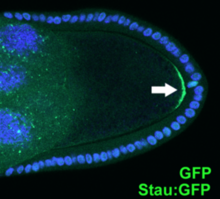 Поляризованная локализация белка Штауфен (белая стрелка) в ооците 9 стадии дрозофилы (Stau: GFP, DAPI).
Поляризованная локализация белка Штауфен (белая стрелка) в ооците 9 стадии дрозофилы (Stau: GFP, DAPI). Эпителиальные клетки прикрепляются друг к другу через плотные соединения, десмосомы и адгезионные соединения, образуя слои клеток, выстилающие поверхность тела животного и внутренние полости (например, пищеварительный тракт и кровеносная система). Эти клетки имеют апикально-базальную полярность, определяемую апикальной мембраной, обращенной к внешней поверхности тела, или просветом внутренних полостей и базолатеральной мембраной ориентированы от просвета. Базолатеральная мембрана относится как к латеральной мембране, где соединения между клетками соединяют соседние клетки, так и к базальной мембране, где клетки прикреплены к базальной мембране, тонкому слою белков внеклеточного матрикса. который отделяет эпителиальный лист от подлежащих клеток и соединительной ткани. Эпителиальные клетки также демонстрируют плоскую клеточную полярность, в которой специализированные структуры ориентированы в плоскости эпителиального слоя. Некоторые примеры плоской полярности клеток включают чешую рыбы, ориентированную в одном направлении, и аналогично перья птиц, мех млекопитающих и кутикулярные выступы (сенсорные волоски и т. Д.) На телах и придатках мух и других насекомых..
Нейрон получает сигналы от соседних клеток через разветвленные клеточные отростки, называемые дендритами. Затем нейрон передает электрический сигнал по специализированному удлинению аксона к синапсу, где высвобождаются нейротрансмиттеры для передачи сигнала другому нейрону или эффекторной клетке (например, мышце или железе). Таким образом, полярность нейрона облегчает направленный поток информации, который необходим для связи между нейронами и эффекторными клетками.
Многие типы клеток могут мигрировать, например лейкоциты и фибробласты, и для того, чтобы эти клетки двигались в одном направлении, они должны иметь определенные переднюю и заднюю части. В передней части клетки находится передний край, который часто определяется плоской волнистостью клеточной мембраны, называемой ламеллиподиумом, или тонкими выступами, называемыми филоподиями. Здесь полимеризация актина в направлении миграции позволяет клеткам расширять передний край клетки и прикрепляться к поверхности. В задней части клетки адгезии разбираются, и пучки актиновых микрофиламентов, называемые стрессорными волокнами, сжимаются и тянут задний край вперед, чтобы не отставать от остальной части клетки. Без этой передне-задней полярности клетки не смогли бы координировать направленную миграцию.
Формирование клеточной полярности критически важно для функционирования практически каждого типа клеток и лежит в основе таких процессов, как деление клеток, дифференциация, миграция клеток, передача сигналов между клетками и оплодотворение. Полярность клеток - это пример свойства самоорганизации, присущего всем живым организмам. Все клетки внутри многоклеточного организма или любого отдельного вида клеток, например дрожжей, демонстрируют поляризованную организацию, необходимую для его пролиферации, дифференцировки или физиологической функции. Бутонированные дрожжи - это очень доступная экспериментальная система, которая служит парадигмой для расшифровки молекулярного механизма, лежащего в основе генерации полярности. Дрожжевые клетки имеют много общих черт в отношении клеточной полярности с другими организмами, например: регулирование внутренними и внешними сигналами, консервативные регуляторные молекулы, такие как Cdc42GTPase, и асимметрия цитоскелета. Поляризация клетки связана с нарушением симметрии, которое происходит в цитоскелете и определяет направление роста будущей дочерней клетки. Это нарушение симметрии облегчает поляризованный поток и локализацию нескольких белков в поляризованном участке. Когда клетки могут выполнять нарушение симметрии в отсутствие какого-либо пространственного ориентира (ориентиры ), это называется спонтанной поляризацией или спонтанным нарушением симметрии.
Короче говоря, установление полярности или нарушение симметрии в данном контексте является первым шагом для определения полярности клеток и, следовательно, деления клеток. Самопроизвольное нарушение симметрии - пример феномена самоорганизации живых клеток.
Молекулярная идентичность наиболее важных белков, участвующих в ведущем спонтанном нарушении симметрии, была тщательно изучена. Эти белки разделены на общие модули, которые представляют собой основные функциональные ядра жизненного цикла дрожжей. Однако молекулярные механизмы, ответственные за эту регуляторную сеть, все еще плохо изучены. Обширная работа, проведенная на многих организмах, от прокариот до высокосложных, выявила центральную роль малых ГТФаз в процессе поляризации клетки. В частности, в дрожжах этим белком является Cdc42, который является членом эукариотического Ras-гомологичного Rho-семейства ГТФаз, которое является частью более широкого суперсемейства малых ГТФаз, включая Rop GTPases в растениях и малые GTPases в прокариотах.
В недавнем исследовании для выяснения связи между временем клеточного цикла и накоплением Cdc42 в зачатке используется оптогенетика для контроля локализации белка с помощью света.
Кроме того, с помощью экспериментальной эволюции были изучены функция и устойчивость установления полярности дрожжей.
Установление полярности у дрожжей обсуждается в Irazoqui and Lew (2004).
Тела позвоночных животных асимметричны по трем осям: передне-задняя (голова к хвосту), дорсально-вентральная (от позвоночника к животу) и слева направо (например, наше сердце находится слева от тела). Эти полярности возникают в развивающемся эмбрионе в результате комбинации нескольких процессов: 1) асимметричное деление клеток, при котором две дочерние клетки получают разное количество клеточного материала (например, мРНК, белков), 2) асимметричная локализация специфических белки или РНК в клетках (которые часто опосредуются цитоскелетом), 3) градиенты концентрации секретируемых белков в эмбрионе, такие как Wnt, Nodal и костные морфогенные белки (BMP), и 4) дифференциальная экспрессия мембранных рецепторов и лигандов, которая вызывает латеральное ингибирование, при котором экспрессирующая рецептор клетка принимает одну судьбу, а ее соседи - другую.
Помимо определения асимметричных осей в во взрослом организме полярность клеток также регулирует как индивидуальные, так и коллективные движения клеток во время эмбрионального развития, такие как апикальное сужение, инвагинация и эпиболия. Эти движения имеют решающее значение для формирования эмбриона и создания сложных структур взрослого тела.
Полярность клеток возникает в основном за счет локализации определенных белков в определенных областях клеточной мембраны. Эта локализация часто требует как рекрутирования цитоплазматических белков на клеточную мембрану, так и поляризованного транспорта везикул вдоль цитоскелетных филаментов для доставки трансмембранных белков из аппарата Гольджи. Многие молекулы, ответственные за регуляцию клеточной полярности, консервативны для разных типов клеток и для всех видов многоклеточных. Примеры включают комплекс PAR (Cdc42, PAR3 / ASIP, PAR6, атипичная протеинкиназа C ), комплекс Крамбса (Crb, PALS, PATJ, Lin7), и комплекс Scribble (Scrib, Dlg, Lgl). Эти комплексы полярности локализованы на цитоплазматической стороне клеточной мембраны асимметрично внутри клеток. Например, в эпителиальных клетках комплексы PAR и Crumbs локализуются вдоль апикальной мембраны, а комплекс Scribble - вдоль боковой мембраны. Вместе с группой сигнальных молекул, называемых Rho GTPases, эти комплексы полярности могут регулировать транспорт везикул, а также контролировать локализацию цитоплазматических белков, главным образом, регулируя фосфорилирование фосфолипидов, называемых фосфоинозитидами. Фосфоинозитиды служат сайтами стыковки белков на клеточной мембране, и их состояние фосфорилирования определяет, какие белки могут связываться.
Хотя многие из белков ключевой полярности хорошо законсервированы, разные механизмы существуют для установления полярности клеток в разных типах клеток. Здесь можно выделить два основных класса: клетки, которые способны спонтанно поляризоваться, и клетки, которые устанавливают полярность на основе внутренних сигналов или сигналов окружающей среды.
Спонтанное нарушение симметрии можно объяснить усилением стохастических флуктуаций молекул из-за отсутствия -линейная химическая кинетика. Математическая основа этого биологического явления была установлена Аланом Тьюрингом в его статье 1953 года «Химические основы морфогенеза ». В то время как Тьюринг первоначально пытался объяснить формирование паттерна в многоклеточной системе, аналогичные механизмы также могут быть применены к формированию внутриклеточного паттерна. Вкратце, если сеть, по крайней мере, из двух взаимодействующих химических веществ (в данном случае белков) демонстрирует определенные типы кинетики реакции, а также дифференциальную диффузию, стохастические флуктуации концентрации могут привести к формированию крупномасштабных стабильных структур, таким образом, переход от от молекулярной длины до клеточного или даже тканевого.
Ярким примером второго типа установления полярности, который зависит от внеклеточных или внутриклеточных сигналов, является зигота C. elegans. Здесь взаимное ингибирование между двумя наборами белков направляет установление и поддержание полярности. С одной стороны, PAR-3, PAR-6 и aPKC (называемые передними белками PAR) занимают как плазматическую мембрану, так и цитоплазму до нарушения симметрии. PAR-1, специфический для C. elegans белок, содержащий безымянный палец PAR-2, и LGL-1 (называемые задними белками PAR) присутствуют в основном в цитоплазме. Мужская центросома обеспечивает сигнал, который нарушает изначально гомогенное мембранное распределение передних PAR, индуцируя кортикальные потоки. Считается, что они адвектируют передние PAR к одной стороне клетки, позволяя задним PAR связываться с другим полюсом (задним). Затем передний и задний белки PAR сохраняют полярность до цитокинеза, взаимно исключая друг друга из соответствующих областей клеточной мембраны.