
Войти
| Сетчатка | |
|---|---|
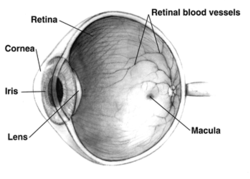 Поперечный разрез правого глаза человека ; глаза значительно различаются у животных. Поперечный разрез правого глаза человека ; глаза значительно различаются у животных. | |
| Подробности | |
| Произношение | Великобритания : / г ɛ т ɪ п ə /, США : / г ɛ т ən ə /, пл. сетчатка / - п я / |
| Часть | Глаз |
| Система | Визуальная система |
| Артерия | Центральная артерия сетчатки |
| Идентификаторы | |
| латинский | Сеть |
| MeSH | D012160 |
| TA98 | A15.2.04.002 |
| TA2 | 6776 |
| FMA | 58301 |
| Анатомическая терминология [ редактировать в Викиданных ] | |
Сетчатка (от латинского : сплетение «чистый») является внутренний, светочувствительный слой ткани глаза большинства позвоночных и некоторых моллюсков. Оптика глаза создает сфокусированное двумерное изображение визуального мира на сетчатке, которое преобразует это изображение в электрические нейронные импульсы в мозг для создания визуального восприятия. Сетчатка выполняет функцию, аналогичную функции пленки или датчика изображения в фотоаппарате.
Нервная сетчатка состоит из нескольких слоев нейронов, связанных между собой синапсами, и поддерживается внешним слоем пигментированных эпителиальных клеток. Первичные светочувствительные клетки сетчатки - это фоторецепторные клетки, которые бывают двух типов: палочки и колбочки. Жезлы работают в основном при тусклом свете и обеспечивают черно-белое изображение. Колбочки работают в хорошо освещенных условиях и отвечают за восприятие цвета, а также за высокую остроту зрения, используемую для таких задач, как чтение. Третий тип светочувствительных клеток, светочувствительные ганглиозные клетки, важен для захвата циркадных ритмов и рефлексивных реакций, таких как зрачковый световой рефлекс.
Свет, падающий на сетчатку, инициирует каскад химических и электрических событий, которые в конечном итоге запускают нервные импульсы, которые отправляются в различные зрительные центры мозга через волокна зрительного нерва. Нейронные сигналы от палочек и колбочек обрабатываются другими нейронами, выход которых принимает форму потенциалов действия в ганглиозных клетках сетчатки, аксоны которых образуют зрительный нерв. Некоторые важные особенности зрительного восприятия можно проследить до кодирования сетчатки глаза и обработки света.
При эмбриональном развитии позвоночных сетчатка и зрительный нерв возникают как выросты развивающегося мозга, особенно эмбрионального промежуточного мозга ; таким образом, сетчатка считается частью центральной нервной системы (ЦНС) и фактически является тканью мозга. Это единственная часть ЦНС, которую можно визуализировать неинвазивно.
Сетчатка позвоночных перевернута в том смысле, что светочувствительные клетки находятся позади сетчатки, так что свет должен проходить через слои нейронов и капилляров, прежде чем достигнет палочек и колбочек. Ганглиозные клетки, аксоны которых образуют зрительный нерв, находятся в передней части сетчатки; поэтому зрительный нерв должен проходить через сетчатку на пути к мозгу. В этой области нет фоторецепторов, что приводит к появлению слепого пятна. Напротив, в сетчатке головоногих фоторецепторы находятся впереди, а обрабатывающие нейроны и капилляры - позади них. Благодаря этому у головоногих моллюсков нет слепого пятна.
Хотя вышележащая нервная ткань частично прозрачна, а сопутствующие глиальные клетки, как было показано, действуют как оптоволоконные каналы для передачи фотонов непосредственно к фоторецепторам, рассеяние света все же происходит. У некоторых позвоночных, в том числе у человека, есть область центральной сетчатки, адаптированная для высокой остроты зрения. Эта область, называемая центральной ямкой, является бессосудистой (не имеет кровеносных сосудов) и имеет минимальную нервную ткань перед фоторецепторами, что сводит к минимуму рассеяние света.
У головоногих моллюсков не перевернутая сетчатка, по разрешающей способности сопоставимая с глазами многих позвоночных. Глаза кальмаров не имеют аналога пигментного эпителия сетчатки позвоночных (РПЭ). Хотя их фоторецепторы содержат белок, ретинохром, который рециркулирует сетчатку и воспроизводит одну из функций РПЭ позвоночных, можно утверждать, что фоторецепторы головоногих моллюсков не поддерживаются так же хорошо, как у позвоночных, и что, в результате, полезный срок службы фоторецепторов в организме беспозвоночные намного короче позвоночных. Легкая замена стебельчатых глаз (у некоторых лобстеров) или сетчатки (у некоторых пауков, таких как Дейнопис) встречается редко.
Сетчатка головоногих не является результатом мозга, как сетчатка позвоночных. Можно утверждать, что это различие показывает, что глаза позвоночных и головоногих не гомологичны, а развивались отдельно. С эволюционной точки зрения более сложная структура, такая как перевернутая сетчатка, обычно может возникать как следствие двух альтернативных процессов: (а) выгодный «хороший» компромисс между конкурирующими функциональными ограничениями или (б) как исторически неадаптивный пережиток прошлого. запутанный путь эволюции и трансформации органов. Зрение - важная адаптация у высших позвоночных.
Третий взгляд на «перевернутый» глаз позвоночных состоит в том, что он сочетает в себе два преимущества: поддержание фоторецепторов, упомянутых выше, и снижение интенсивности света, необходимое для предотвращения ослепления фоторецепторов, которые основаны на чрезвычайно чувствительных глазах предков современные миксы (рыба, обитающая в очень глубокой темной воде).
 Раздел сетчатки
Раздел сетчатки  Палочки, колбочки и нервные слои сетчатки. Передняя (передняя) часть глаза находится слева. Свет (слева) проходит через несколько прозрачных нервных слоев и достигает стержней и колбочек (крайний справа). Химические изменения в палочках и колбочках посылают сигнал обратно к нервам. Сигнал сначала поступает в биполярные и горизонтальные клетки (желтый слой), затем в амакриновые клетки и ганглиозные клетки (фиолетовый слой), а затем в волокна зрительного нерва. В этих слоях обрабатываются сигналы. Во-первых, сигналы начинаются как необработанные выходные данные точек в ячейках стержня и конуса. Затем нервные слои идентифицируют простые формы, такие как яркие точки в окружении темных точек, края и движения. (На основе рисунка Рамона-и-Кахала, 1911 г.)
Палочки, колбочки и нервные слои сетчатки. Передняя (передняя) часть глаза находится слева. Свет (слева) проходит через несколько прозрачных нервных слоев и достигает стержней и колбочек (крайний справа). Химические изменения в палочках и колбочках посылают сигнал обратно к нервам. Сигнал сначала поступает в биполярные и горизонтальные клетки (желтый слой), затем в амакриновые клетки и ганглиозные клетки (фиолетовый слой), а затем в волокна зрительного нерва. В этих слоях обрабатываются сигналы. Во-первых, сигналы начинаются как необработанные выходные данные точек в ячейках стержня и конуса. Затем нервные слои идентифицируют простые формы, такие как яркие точки в окружении темных точек, края и движения. (На основе рисунка Рамона-и-Кахала, 1911 г.)  Иллюстрация распределения колбочек в ямке человека с нормальным цветовым зрением (слева) и сетчаткой с дальтонизмом (протанопией). Обратите внимание, что в центре ямки находится очень мало колбочек, чувствительных к синему.
Иллюстрация распределения колбочек в ямке человека с нормальным цветовым зрением (слева) и сетчаткой с дальтонизмом (протанопией). Обратите внимание, что в центре ямки находится очень мало колбочек, чувствительных к синему.  Распределение палочек и колбочек по линии, проходящей через ямку и слепое пятно человеческого глаза
Распределение палочек и колбочек по линии, проходящей через ямку и слепое пятно человеческого глаза Сетчатка позвоночных состоит из десяти отдельных слоев. От самого близкого до самого дальнего от стекловидного тела:
Эти слои можно сгруппировать в 4 основных этапа обработки: фоторецепция; передача биполярным клеткам ; передача ганглиозным клеткам, которые также содержат фоторецепторы, светочувствительным ганглиозным клеткам ; и передача по зрительному нерву. На каждой синаптической стадии также присутствуют латерально соединяющиеся горизонтальные и амакриновые клетки.
Зрительный нерв является одним из центральных путей многих аксонов ганглиозных клеток, соединяющих прежде всего к боковому коленчатому тела, визуальная ретрансляционной станции в диэнцефалона ( в заднюю части переднего мозга). Он также проецируется на верхний бугорок, супрахиазматическое ядро и ядро зрительного тракта. Он проходит через другие слои, образуя диск зрительного нерва у приматов.
Дополнительные структуры, не связанные напрямую со зрением, обнаруживаются в виде выростов сетчатки у некоторых групп позвоночных. В птицах, то гребень является сосудистой структурой сложной формы, что выступает из сетчатки в стекловидное тело ; он снабжает глаза кислородом и питательными веществами, а также может помочь зрению. Рептилии имеют похожее, но гораздо более простое строение.
У взрослых людей вся сетчатка составляет примерно 72% сферы диаметром около 22 мм. Вся сетчатка содержит около 7 миллионов колбочек и от 75 до 150 миллионов палочек. Диск зрительного нерва, часть сетчатки, которую иногда называют «слепым пятном» из-за отсутствия фоторецепторов, расположен в зрительном сосочке, где волокна зрительного нерва выходят из глаза. Он выглядит как овальная белая область площадью 3 мм². Височная (в направлении висков) к этому диску находится макула, в центре которой находится ямка, ямка, которая отвечает за наше резкое центральное зрение, но на самом деле менее чувствительна к свету из-за отсутствия стержней. У человека и нечеловеческих приматов есть одна ямка, в отличие от некоторых видов птиц, таких как ястребы, которые имеют двояковыпуклый вид, а также собаки и кошки, у которых нет ямки, но есть центральная полоса, известная как зрительная полоса. Вокруг ямки проходит центральная сетчатка примерно на 6 мм, а затем и периферическая сетчатка. Самый дальний край сетчатки определяется зубчатой линией. Расстояние от одного или другого до другого (или макулы), наиболее чувствительной области вдоль горизонтального меридиана, составляет около 32 мм.
На срезе сетчатка не более 0,5 мм толщиной. Он состоит из трех слоев нервных клеток и двух синапсов, включая уникальный ленточный синапс. Зрительный нерв переносит аксоны ганглиозных клеток в мозг и кровеносные сосуды, снабжающие сетчатку. Ганглиозные клетки лежат внутри глаза, а светочувствительные клетки - за его пределами. Из-за такого противоречивого расположения свет должен сначала пройти через ганглиозные клетки и через толщу сетчатки (включая ее капиллярные сосуды, не показаны), прежде чем достигнет палочек и колбочек. Свет поглощается пигментным эпителием сетчатки или сосудистой оболочкой (оба непрозрачны).
В лейкоциты в капиллярах перед фоторецепторов может восприниматься как небольшие яркие движущихся точек при взгляде на синий свет. Это известно как энтоптический феномен синего поля (или феномен Шерера ).
Между слоем ганглиозных клеток и палочками и колбочками находятся два слоя нейропилей, в которых осуществляются синаптические контакты. Слои нейропиля - это внешний плексиформный слой и внутренний плексиформный слой. Во внешнем слое нейропиля палочки и колбочки соединяются с вертикально движущимися биполярными клетками, а горизонтально ориентированные горизонтальные клетки соединяются с ганглиозными клетками.
Центральная сетчатка преимущественно содержит колбочки, а периферическая сетчатка - преимущественно палочки. Всего насчитывается около семи миллионов колбочек и сто миллионов стержней. В центре макулы находится фовеальная ямка, в которой колбочки узкие и длинные и расположены в виде шестиугольной мозаики, наиболее плотной, в отличие от гораздо более толстых колбочек, расположенных на периферии сетчатки. В фовеальной ямке смещаются другие слои сетчатки перед тем, как нарастать вдоль склона фовеа до тех пор, пока не будет достигнут край фовеа, или парафовеа, который является самой толстой частью сетчатки. Макула имеет желтую пигментацию из-за скрининговых пигментов и известна как желтое пятно. Область, непосредственно окружающая фовеа, имеет самую высокую плотность палочек, сходящихся на единичных биполярных клетках. Поскольку ее конусы имеют гораздо меньшую сходимость сигналов, ямка обеспечивает самое резкое зрение, которое только может получить глаз.
Хотя палочка и колбочки представляют собой своего рода мозаику, передача от рецепторов к биполярам и ганглиозным клеткам не является прямой. Поскольку существует около 150 миллионов рецепторов и только 1 миллион волокон зрительного нерва, должна происходить конвергенция и, следовательно, смешение сигналов. Более того, горизонтальное действие горизонтальных и амакриновых клеток может позволить одной области сетчатки контролировать другую (например, один стимул подавляет другой). Это торможение является ключом к уменьшению количества сообщений, отправляемых в высшие области мозга. У некоторых низших позвоночных (например, у голубей ) существует «центробежный» контроль сообщений - то есть один слой может контролировать другой, или более высокие области мозга могут управлять нервными клетками сетчатки, но у приматов этого не происходит.
С помощью оптической когерентной томографии (ОКТ) в сетчатке можно идентифицировать 18 слоев. Слои и анатомическая корреляция следующие:
 ОКТ во временной области макулярной области сетчатки при 800 нм, аксиальное разрешение 3 мкм
ОКТ во временной области макулярной области сетчатки при 800 нм, аксиальное разрешение 3 мкм 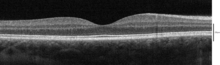 ОКТ-сканирование поперечного сечения макулы в спектральной области.
ОКТ-сканирование поперечного сечения макулы в спектральной области.  гистология макулы (ОКТ)
гистология макулы (ОКТ) От самого внутреннего до самого внешнего, уровни, идентифицируемые OCT, следующие:
| # | Слой OCT / Обычная этикетка | Анатомический коррелят | Отражательная способность | Специфический анатомический границы? | Дополнительный использованная литература |
|---|---|---|---|---|---|
| 1 | Задний кортикальный слой стекловидного тела | Задний кортикальный слой стекловидного тела | Гиперотражающий | да | |
| 2 | Преретинальное пространство | В глазах, где стекловидное тело полностью или частично отделилось от сетчатки, это пространство, созданное между задней кортикальной стороной стекловидного тела и внутренней ограничивающей мембраной сетчатки. | Гипоотражающий | ||
| 3 | Внутренняя ограничивающая мембрана (ILM) | Сформированные клетки Мюллера endfeet (неясно, наблюдается ли это на ОКТ) | Гиперотражающий | Нет | |
| Слой нервных волокон (NFL) | Аксоны ганглиозных клеток движутся к зрительному нерву | ||||
| 4 | Слой ганглиозных клеток (GCL) | Тела ганглиозных клеток (и некоторые смещенные амакриновые клетки ) | Гипоотражающий | ||
| 5 | Внутренний плексиформный слой (IPL) | Синапсы между биполярными, амакриновыми и ганглиозными клетками | Гиперотражающий | ||
| 6 | Внутренний ядерный слой (INL) | а) Горизонтальные, биполярные и амакриновые клеточные тела б) Ядра клеток Мюллера | Гипоотражающий | ||
| 7 | Внешний плексиформный слой (OPL) | Синапсы между фоторецепторами, биполярными и горизонтальными клетками | Гиперотражающий | ||
| 8 | (Внутренняя половина) слой нервных волокон Генле (HL) | Фоторецепторные аксоны (косо ориентированные волокна; отсутствуют в средней или периферической сетчатке) | Гипоотражающий | Нет | |
| (Внешняя половина) Внешний ядерный слой (ONL) | В фоторецепторах клеточных тел | ||||
| 9 | Внешняя ограничивающая мембрана (ELM) | Изготовлен из зонул, прикрепленных между клетками Мюллера и внутренними сегментами фоторецепторов. | Гиперотражающий | ||
| 10 | Миоидная зона (МЗ) | Самая внутренняя часть внутреннего сегмента фоторецептора (IS), содержащая:
| Гипоотражающий | Нет | |
| 11 | Зона эллипсоида (EZ) | Самая удаленная часть внутреннего сегмента фоторецептора (IS) упакована митохондриями. | Очень гиперотражающий | Нет | |
| Соединение IS / OS или линия целостности фоторецепторов (PIL) | Фоторецептор, соединяющий реснички, соединяющие внутренний и внешний сегменты фоторецепторных клеток. | ||||
| 12 | Наружные сегменты фоторецепторов (OS) | На наружных сегментов фоторецепторов (ОС), которые содержат диски, заполненные опсина, молекула, которая поглощает фотоны. | Гипоотражающий | ||
| 13 | Зона пересечения (IZ) | Вершины клеток РПЭ, покрывающие часть ОС колбочек. Слабо отличим от РПЭ. Ранее: «Линия вершин внешнего сегмента конуса» (СТОИМОСТЬ) | Гиперотражающий | Нет | |
| 14 | Комплекс РПЭ / Бруха | Фагосомная зона РПЭ | Очень гиперотражающий | Нет | |
| Зона меланосом РПЭ | Гипоотражающий | ||||
| Зона митохондрий RPE + соединение между RPE и мембраной Бруха | Очень гиперотражающий | ||||
| 15 | Хориокапилляры | Тонкий слой умеренной отражательной способности во внутренней сосудистой оболочке | Нет | ||
| 16 | Слой Саттлера | Толстый слой гиперотражающих профилей круглой или овальной формы с гипорефлективными сердцевинами в средней части сосудистой оболочки. | |||
| 17 | Слой Галлера | Толстый слой овальных гиперотражающих профилей с гипорефлективными сердцевинами в наружной сосудистой оболочке | |||
| 18 | Хориоидально-склеральный переход | Зона внешней хориоидеи с заметным изменением текстуры, в которой большие круглые или яйцевидные профили примыкают к однородная область переменной отражательной способности | |||
Развитие сетчатки начинается с образования глазных полей, опосредованных белками SHH и SIX3, с последующим развитием зрительных пузырьков, регулируемых белками PAX6 и LHX2. Роль Pax6 в развитии глаз была элегантно продемонстрирована Walter Gehring с коллегами, которые показали, что эктопическая экспрессия Pax6 может приводить к формированию глаз на антеннах, крыльях и ногах дрозофилы. Оптический пузырек дает начало трем структурам: нервной сетчатке, пигментированному эпителию сетчатки и оптическому стеблю. Нервная сетчатка содержит клетки-предшественники сетчатки (RPC), которые дают начало семи типам клеток сетчатки. Дифференциация начинается с ганглиозных клеток сетчатки и завершается образованием глии Мюллера. Хотя каждый тип клеток дифференцируется от RPC в последовательном порядке, существует значительное перекрытие во времени, когда дифференцируются отдельные типы клеток. Сигналы, которые определяют судьбу дочерних клеток RPC, кодируются множеством семейств факторов транскрипции, включая факторы bHLH и гомеодомена.
Помимо определения судьбы направляющих клеток, в сетчатке существуют сигналы для определения дорсально-вентральной (DV) и назально-височной (NT) осей. Ось DV устанавливается с помощью вентрального к дорсальному градиенту VAX2, тогда как ось NT координируется экспрессией факторов транскрипции вилки FOXD1 и FOXG1. Внутри сетчатки образуются дополнительные градиенты. Это пространственное распределение может помочь в правильном нацеливании на аксоны RGC, которые функционируют, чтобы установить ретинотопную карту.
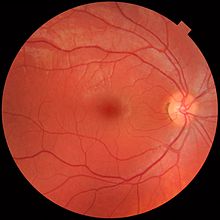 Фотография глазного дна, показывающая кровеносные сосуды в сетчатке нормального человека. Вены темнее и немного шире соответствующих артерий. Оптический диск находится справа, а желтое пятно находится недалеко от центра.
Фотография глазного дна, показывающая кровеносные сосуды в сетчатке нормального человека. Вены темнее и немного шире соответствующих артерий. Оптический диск находится справа, а желтое пятно находится недалеко от центра. Сетчатка разделена на отдельные слои, каждый из которых содержит определенные типы клеток или клеточные компартменты, метаболизм которых имеет различные потребности в питании. Чтобы удовлетворить эти требования, глазная артерия разветвляется и снабжает сетчатку двумя различными сосудистыми сетями: хориоидальной сетью, которая снабжает сосудистую оболочку и внешнюю сетчатку, и сетчаткой сетчаткой, которая снабжает внутренний слой сетчатки.
На первый взгляд, можно подумать, что сетчатка позвоночного «неправильно спроектирована» или «плохо спроектирована»; но на самом деле сетчатка не могла бы функционировать, если бы ее не перевернули. Слой фоторецепторов должен быть встроен в пигментный эпителий сетчатки (ППЭ), который выполняет по крайней мере семь жизненно важных функций, одна из наиболее очевидных - это снабжение кислородом и другими необходимыми питательными веществами, необходимыми для функционирования фоторецепторов. Эти питательные вещества включают глюкозу, жирные кислоты и сетчатку. Процесс амплификации фоторецепторов млекопитающих использует большие количества энергии для зрения в фотопических условиях (требуя меньше в скотопических условиях) и, таким образом, требует большого количества питательных веществ, поставляемых кровеносными сосудами в сосудистой оболочке, которая находится за пределами РПЭ. Сосудистая оболочка снабжает сетчатку около 75% этих питательных веществ, а сосудистая сеть сетчатки - только 25%.
Когда свет попадает на 11-цис-ретиналь (диски в палочках и колбочках), 11-цис-сетчатка превращается в полностью транс-ретиналь, что затем вызывает изменения в опсинах. Теперь внешние сегменты не регенерируют сетчатку обратно в цис-форму после того, как она была изменена светом. Вместо этого сетчатка откачивается к окружающему РПЭ, где она регенерируется и транспортируется обратно во внешние сегменты фоторецепторов. Эта функция рециркуляции RPE защищает фоторецепторы от фотоокислительного повреждения и позволяет фоторецепторным клеткам иметь полезный срок службы на протяжении десятилетий.
Сетчатка птицы лишена кровеносных сосудов, возможно, для обеспечения беспрепятственного прохождения света для формирования изображений, что дает лучшее разрешение. Следовательно, считается, что сетчатка птицы питается и снабжается кислородом от специализированного органа, называемого «pecten» или pecten oculi, расположенного в слепом пятне или на диске зрительного нерва. Этот орган чрезвычайно богат кровеносными сосудами и, как считается, снабжает сетчатку птиц питанием и кислородом путем диффузии через стекловидное тело. Пектен очень богат активностью щелочной фосфатазы и поляризованными клетками в его мостиковой части - и то и другое соответствует его секреторной роли. Клетки пектена заполнены темными гранулами меланина, которые, как предполагалось, сохраняют этот орган в тепле за счет поглощения паразитного света, падающего на пектен. Считается, что это увеличивает скорость метаболизма пектена, тем самым экспортируя больше питательных молекул для удовлетворения строгих энергетических потребностей сетчатки в течение длительных периодов воздействия света.
Известно, что бифуркации и другие физические характеристики внутренней сосудистой сети сетчатки у разных людей различаются, и эти индивидуальные различия использовались для биометрической идентификации и для раннего выявления начала заболевания. Картирование сосудистых бифуркаций - один из основных этапов биометрической идентификации. Результаты такого анализа структуры кровеносных сосудов сетчатки могут быть оценены на основе достоверных данных о сосудистых бифуркациях изображений глазного дна сетчатки, полученных из набора данных DRIVE. Кроме того, были определены классы судов из набора данных DRIVE, и также доступен автоматизированный метод для точного выделения этих бифуркаций. Изменения в кровообращении сетчатки наблюдаются с возрастом и воздействием загрязненного воздуха и могут указывать на сердечно-сосудистые заболевания, такие как гипертония и атеросклероз. Определение эквивалентной ширины артериол и венул около диска зрительного нерва также является широко используемым методом выявления сердечно-сосудистых рисков.
Сетчатка преобразует оптическое изображение в нервные импульсы, начиная с шаблонного возбуждения цветочувствительных пигментов своих палочек и колбочек, фоторецепторных клеток сетчатки. Возбуждение обрабатывается нейронной системой и различными частями мозга, работающими параллельно, чтобы сформировать представление о внешней среде в мозгу.
Колбочки реагируют на яркий свет и обеспечивают цветовое зрение с высоким разрешением при дневном освещении (также называемое фотопическим зрением ). Реакции палочек насыщены при дневном свете и не влияют на видение паттернов. Однако стержни действительно реагируют на тусклый свет и обеспечивают монохроматическое зрение с низким разрешением при очень низких уровнях освещения (так называемое скотопическое зрение ). Освещение в большинстве офисных помещений находится между этими двумя уровнями и называется мезопическим зрением. На мезопических уровнях освещения и стержни, и колбочки активно передают информацию о структуре. Какой вклад информация о стержне вносит в видение паттернов в этих условиях, неясно.
Реакция колбочек на световые волны различной длины называется их спектральной чувствительностью. В нормальном человеческом зрении спектральная чувствительность конуса подразделяется на один из трех подтипов, часто называемых синим, зеленым и красным, но более точно известный как подтипы колбочек, чувствительных к короткой, средней и длинной волнам. Отсутствие одного или нескольких подтипов колбочек приводит к тому, что у людей наблюдается дефицит цветового зрения или различные виды дальтонизма. Эти люди не слепы к предметам определенного цвета, но не могут различать цвета, которые могут различить люди с нормальным зрением. У людей есть это трехцветное зрение, в то время как у большинства других млекопитающих нет колбочек с чувствительным к красному пигментом, и поэтому у них плохое двухцветное цветовое зрение. Однако у некоторых животных есть четыре спектральных подтипа, например, форель добавляет ультрафиолетовую подгруппу к коротким, средним и длинным подтипам, которые похожи на людей. Некоторые рыбы также чувствительны к поляризации света.
В фоторецепторах воздействие света гиперполяризует мембрану в виде серии ступенчатых сдвигов. Внешний сегмент клетки содержит фотопигмент. Внутри клетки нормальный уровень циклического гуанозинмонофосфата (цГМФ) сохраняет канал Na + открытым, и, таким образом, в состоянии покоя клетка деполяризуется. Фотона приводит к тому, ретиналь, связанный с белком рецептора к изомеризации с транс-ретиналя. Это заставляет рецептор активировать несколько G-белков. Это, в свою очередь, заставляет Ga-субъединицу белка активировать фосфодиэстеразу (PDE6), которая разрушает цГМФ, что приводит к закрытию ионных каналов, управляемых циклическими нуклеотидами (CNG) Na +. Таким образом, клетка гиперполяризована. Количество выделяемого нейротрансмиттера уменьшается при ярком свете и увеличивается с понижением уровня освещенности. Настоящий фотопигмент обесцвечивается при ярком свете и заменяется только в результате химического процесса, поэтому при переходе от яркого света к темноте глазу может потребоваться до тридцати минут, чтобы достичь полной чувствительности.
Когда, таким образом, возбуждаются светом, photoceptor посылает пропорциональный ответ синаптический на биполярные клетки, которые, в своей очереди сигнализируют ганглиозные клетки сетчатки. Фоторецепторы также сшиты горизонтальными клетками и амакриновыми клетками, которые изменяют синаптический сигнал до того, как он достигает ганглиозных клеток, при этом нервные сигналы перемешиваются и комбинируются. Из нервных клеток сетчатки только ганглиозные клетки сетчатки и несколько амакриновых клеток создают потенциалы действия.
В ганглиозных клетках сетчатки есть два типа ответа, в зависимости от рецептивного поля клетки. Воспринимающие поля ганглиозных клеток сетчатки включают центральную, приблизительно круглую область, где свет оказывает одно влияние на активацию клетки, и кольцевое окружение, где свет имеет противоположный эффект. В ячейках ON увеличение интенсивности света в центре воспринимающего поля приводит к увеличению скорости стрельбы. В ВЫКЛЮЧЕННЫХ ячейках заставляет его уменьшаться. В линейной модели этот профиль отклика хорошо описывается разностью гауссианов и является основой для алгоритмов обнаружения границ. Помимо этого простого различия, ганглиозные клетки также различаются по хроматической чувствительности и типу пространственного суммирования. Клетки, демонстрирующие линейное пространственное суммирование, называются X-клетками (также называемыми парвоцеллюлярными, P или сверхмалыми ганглиозными клетками), а те, которые демонстрируют нелинейное суммирование, являются Y-клетками (также называемыми магноклеточными, M или паразольными ганглиозными клетками сетчатки), хотя соответствие между X- и Y-клетки (в сетчатке кошки) и P- и M-клетки (в сетчатке приматов) не так просты, как когда-то казалось.
При передаче зрительных сигналов в мозг, зрительный путь, сетчатка по вертикали делится на две части: височную (ближе к виску) и носовую (ближе к носу) половину. Аксоны из носовой половины пересекают мозг через перекрест зрительных нервов, чтобы соединиться с аксонами из височной половины другого глаза, прежде чем перейти в латеральное коленчатое тело.
Хотя существует более 130 миллионов рецепторов сетчатки, в зрительном нерве насчитывается только около 1,2 миллиона волокон (аксонов). Таким образом, большая часть предварительной обработки выполняется внутри сетчатки. Ямка производит наиболее точную информацию. Несмотря на то, что они занимают около 0,01% поля зрения (менее 2 ° угла зрения ), около 10% аксонов зрительного нерва посвящены ямке. Предел разрешения фовеа составляет около 10 000 точек. Информационная емкость оценивается в 500 000 бит в секунду (для получения дополнительной информации о битах см. Теорию информации ) без цвета или около 600 000 бит в секунду, включая цвет.
 В центре и вне центра сетчатки
В центре и вне центра сетчатки Когда сетчатка посылает в мозг нейронные импульсы, представляющие изображение, она пространственно кодирует (сжимает) эти импульсы, чтобы соответствовать ограниченной емкости зрительного нерва. Сжатие необходимо, потому что фоторецепторных клеток в 100 раз больше, чем ганглиозных клеток. Это осуществляется « декорреляцией », которая осуществляется «структурами центр-окружение», которые реализуются биполярными и ганглиозными клетками.
В сетчатке есть два типа центрально-окружающих структур - на центрах и вне центра. На центрах есть положительно взвешенный центр и отрицательно взвешенный объемный звук. Внецентры - как раз наоборот. Положительное взвешивание более известно как возбуждающее, а отрицательное - как тормозящее.
Эти центрально-окружающие структуры не являются физически очевидными в том смысле, что их нельзя увидеть, окрашивая образцы ткани и исследуя анатомию сетчатки. Структуры центр-окружение логичны (т.е. математически абстрактны) в том смысле, что они зависят от силы связи между биполярными и ганглиозными клетками. Считается, что сила связи между клетками обусловлена количеством и типами ионных каналов, встроенных в синапсы между биполярными и ганглиозными клетками.
Структуры центр-окружение математически эквивалентны алгоритмам обнаружения краев, используемым компьютерными программистами для выделения или улучшения краев цифровой фотографии. Таким образом, сетчатка выполняет операции с импульсами, представляющими изображение, для улучшения краев объектов в пределах своего поля зрения. Например, на изображении собаки, кошки и машины именно края этих объектов содержат больше всего информации. Чтобы высшие функции мозга (или компьютера) извлекали и классифицировали такие объекты, как собака и кошка, сетчатка является первым шагом к разделению различных объектов в пределах сцены.
Например, следующая матрица лежит в основе компьютерного алгоритма, реализующего обнаружение границ. Эта матрица является компьютерным эквивалентом структуры центр-окружение. В этом примере каждый блок (элемент) в этой матрице будет связан с одним фоторецептором. Фоторецептор в центре - это рецептор тока, который обрабатывается. Центральный фоторецептор умножается на весовой коэффициент +1. Окружающие фоторецепторы являются «ближайшими соседями» к центру и умножаются на значение -1/8. Окончательно подсчитывается сумма всех девяти этих элементов. Это суммирование повторяется для каждого фоторецептора в изображении путем сдвига влево до конца строки, а затем вниз до следующей строки.
| -1/8 | -1/8 | -1/8 |
| -1/8 | +1 | -1/8 |
| -1/8 | -1/8 | -1/8 |
Общая сумма этой матрицы равна нулю, если все входы от девяти фоторецепторов имеют одинаковое значение. Нулевой результат означает, что изображение было однородным (неизменным) в пределах этого небольшого участка. Отрицательные или положительные суммы означают, что изображение изменялось (изменялось) в пределах этого небольшого участка из девяти фоторецепторов.
Вышеупомянутая матрица является лишь приближением того, что на самом деле происходит внутри сетчатки. Отличия заключаются в следующем:
Вот пример входного изображения и того, как обнаружение краев изменит его.

Как только изображение пространственно кодируется структурами центр-окружение, сигнал посылается по зрительному нерву (через аксоны ганглиозных клеток) через зрительный перекрест в LGN ( латеральное коленчатое ядро ). Точная функция LGN в настоящее время неизвестна. Затем результат LGN отправляется в заднюю часть мозга. В частности, выход LGN «излучается» в первичную зрительную кору V1.
Упрощенный поток сигнала: фоторецепторы → биполярный → ганглиоз → хиазма → LGN → кора V1.

Существует множество наследственных и приобретенных заболеваний или нарушений, которые могут повлиять на сетчатку. Некоторые из них включают:
Доступен ряд различных инструментов для диагностики заболеваний и нарушений, влияющих на сетчатку. Офтальмоскопия и фотография глазного дна давно используются для исследования сетчатки. Недавно адаптивная оптика использовалась для изображения отдельных палочек и колбочек в сетчатке живого человека, а компания из Шотландии разработала технологию, которая позволяет врачам наблюдать сетчатку целиком без какого-либо дискомфорта для пациентов.
Электроретинограмма используется для неинвазивного измерения электрической активности сетчатки, на которую влияют некоторыми заболевания. Относительно новой технологией, которая сейчас становится широко доступной, является оптическая когерентная томография (ОКТ). Этот неинвазивный метод позволяет получить объемную трехмерную томограмму или томограмму поперечного сечения тонких структур сетчатки с высоким разрешением с гистологическим качеством. Анализ сосудов сетчатки - это неинвазивный метод исследования мелких артерий и вен сетчатки, который позволяет сделать выводы о морфологии и функции мелких сосудов в других частях человеческого тела. Было установлено, что он является предиктором сердечно-сосудистых заболеваний и, согласно исследованию, опубликованному в 2019 году, имеет потенциал для раннего выявления болезни Альцгеймера.
Лечение зависит от характера заболевания или расстройства.
Ниже приведены общие методы лечения заболеваний сетчатки:
| Редкие или нетипичные методы лечения заболеваний сетчатки |
|---|
| Генная терапия сетчатки Основная статья: Аденоассоциированный вирус и генная терапия сетчатки глаза человекаГенная терапия обещает стать потенциальным средством лечения широкого спектра заболеваний сетчатки. Это включает использование неинфекционного вируса для переноса гена в часть сетчатки. Векторы рекомбинантного аденоассоциированного вируса (rAAV) обладают рядом особенностей, которые делают их идеально подходящими для генной терапии сетчатки, включая отсутствие патогенности, минимальную иммуногенность и способность стабильно и эффективно преобразовывать постмитотические клетки. Векторы rAAV все чаще используются из-за их способности опосредовать эффективную трансдукцию пигментного эпителия сетчатки (RPE), фоторецепторных клеток и ганглиозных клеток сетчатки. Каждый тип клеток может быть целенаправленным путем выбора соответствующей комбинации серотипа AAV, промотора и места внутриглазной инъекции. Несколько клинических испытаний уже сообщили о положительных результатах использования rAAV для лечения врожденного амавроза Лебера, показав, что терапия была безопасной и эффективной. Серьезных побочных эффектов не было, и пациенты во всех трех исследованиях показали улучшение зрительной функции, измеренное рядом методов. Используемые методы варьировались в трех испытаниях, но включали как функциональные методы, такие как острота зрения и функциональная подвижность, так и объективные измерения, которые менее подвержены смещению, такие как способность зрачка реагировать на свет и улучшения функциональной МРТ. Улучшение сохранялось в течение длительного времени, и пациенты продолжали чувствовать себя хорошо спустя более 1,5 лет. Этому процессу способствует уникальная архитектура сетчатки и ее относительно защищенная иммунитетом среда. Плотные соединения, которые образуют гемато-ретинальный барьер, отделяют субретинальное пространство от кровоснабжения, таким образом защищая его от микробов и большинства иммуно-опосредованных повреждений и повышая его способность реагировать на векторно-опосредованную терапию. Анатомия глаза с высокой степенью компартментализации способствует точной доставке суспензий терапевтических векторов к конкретным тканям при прямой визуализации с использованием микрохирургических методов. В защищенной среде сетчатки векторы AAV способны поддерживать высокие уровни экспрессии трансгена в пигментированном эпителии сетчатки (RPE), фоторецепторах или ганглиозных клетках в течение длительных периодов времени после однократной обработки. Кроме того, глаз и зрительную систему можно регулярно и легко контролировать на предмет зрительной функции и структурных изменений сетчатки после инъекций с помощью неинвазивных передовых технологий, таких как острота зрения, контрастная чувствительность, автофлуоресценция глазного дна (FAF), адаптированные к темноте пороги зрения., диаметры сосудов, пупиллометрия, электроретинография (ЭРГ), мультифокальная ЭРГ и оптическая когерентная томография (ОКТ). Эта стратегия эффективна против ряда изученных заболеваний сетчатки, включая неоваскулярные заболевания, которые являются признаками возрастной дегенерации желтого пятна, диабетической ретинопатии и ретинопатии недоношенных. Поскольку регуляция васкуляризации в зрелой сетчатке включает баланс между эндогенными положительными факторами роста, такими как фактор роста эндотелия сосудов (VEGF), и ингибиторами ангиогенеза, такими как фактор, происходящий из пигментного эпителия ( PEDF ), rAAV-опосредованная экспрессия PEDF, ангиостатин и растворимый рецептор VEGF sFlt-1, которые все являются антиангиогенными белками, уменьшают образование аберрантных сосудов на животных моделях. Поскольку специфические генные методы лечения не могут быть легко использованы для лечения значительной части пациентов с дистрофией сетчатки, существует большой интерес к разработке более широко применимой терапии факторами выживаемости. Нейротрофические факторы обладают способностью модулировать рост нейронов во время развития, чтобы поддерживать существующие клетки и обеспечивать восстановление поврежденных популяций нейронов в глазу. AAV, кодирующие нейротрофические факторы, такие как члены семейства факторов роста фибробластов (FGF) и GDNF, либо защищали фоторецепторы от апоптоза, либо замедляли гибель клеток. Орган Трансплантация Трансплантация сетчаток была предпринята попытка, но без особого успеха. В Массачусетском технологическом институте, Университете Южной Калифорнии, Университете Аахена и Университете Нового Южного Уэльса разрабатывается «искусственная сетчатка»: имплант, который будет обходить фоторецепторы сетчатки и напрямую стимулировать прикрепленные нервные клетки с помощью сигналов. с цифровой камеры. |
Около 300 г. до н.э., Герофил определил сетчатку от рассечений трупных глаз. Он назвал его паутинным слоем из-за его сходства с паутиной и retiform из-за его сходства с сетью для забросов. Термин паутинная оболочка стал обозначать слой вокруг мозга; термин retiform стал обозначать сетчатку.
Между 1011 и 1021 годами нашей эры Ибн аль-Хайтам опубликовал многочисленные эксперименты, демонстрирующие, что зрение возникает из света, отражающегося от объектов в глаза. Это согласуется с теорией внутреннего деления и против теории излучения, теории, согласно которой зрение происходит от лучей, испускаемых глазами. Однако Ибн аль-Хайтам решил, что сетчатка не может быть ответственна за начало зрения, потому что изображение, сформированное на ней, было перевернуто. Вместо этого он решил, что все должно начинаться с поверхности линзы.
В 1604 году Иоганн Кеплер разработал оптику глаза и решил, что сетчатка должна быть там, где начинается зрение. Он предоставил другим ученым возможность согласовать перевернутое изображение на сетчатке глаза с нашим восприятием мира как вертикального.
В 1894 году Сантьяго Рамон-и-Кахаль опубликовал первую важную характеристику нейронов сетчатки в Retina der Wirbelthiere ( Сетчатка позвоночных).
Джордж Уолд, Халдан Кеффер Хартлайн и Рагнар Гранит получили Нобелевскую премию по физиологии и медицине 1967 года за свои научные исследования сетчатки.
Недавнее исследование Пенсильванского университета подсчитало, что приблизительная пропускная способность сетчатки глаза человека составляет 8,75 мегабит в секунду, тогда как скорость передачи данных через сетчатку морской свинки составляет 875 килобит в секунду.
Макларен и Пирсон и его коллеги из Университетского колледжа Лондона и глазной больницы Мурфилдс в Лондоне в 2006 году показали, что фоторецепторные клетки могут быть успешно трансплантированы в сетчатку мыши, если донорские клетки находятся на критической стадии развития. Недавно Адер и его коллеги в Дублине показали с помощью электронного микроскопа, что трансплантированные фоторецепторы образуют синаптические связи.
В 2012 году Себастьян Сын и его лаборатория в Массачусетском технологическом институте запустили EyeWire, онлайн- игру Citizen, в которой игроки отслеживают нейроны сетчатки. Цели проекта EyeWire - выявить определенные типы клеток в пределах известных широких классов клеток сетчатки и составить карту связей между нейронами сетчатки, что поможет определить, как работает зрение.

Структуры глаза обозначены

Другой вид глаза и структуры глаза с пометкой

Иллюстрация изображения, «видимого» сетчаткой независимо от обработки зрительного нерва и полосатой коры.