
Войти
 Раздел сетчатки: свет сначала падает на слой ганглиозных клеток, последние палочки и колбочки
Раздел сетчатки: свет сначала падает на слой ганглиозных клеток, последние палочки и колбочки по своей природе светочувствительные ганглиозные клетки сетчатки (ipRGC ), также называемые светочувствительными ганглиозными клетками сетчатки (pRGC ), или содержащие меланопсин ганглиозные клетки сетчатки (mRGC ) представляют собой тип нейрона в сетчатке глаза млекопитающего. Присутствие ipRGC было впервые отмечено в 1923 году, когда мыши без палочек и конусов все еще отвечали на световой стимул сужением зрачка, что свидетельствует о том, что палочки и колбочки - не единственные светочувствительные нейроны сетчатки. Только в 1980-х годах начались исследования этих клеток. Недавние исследования показали, что эти ганглиозные клетки сетчатки, в отличие от других ганглиозных клеток сетчатки, по своей природе светочувствительны из-за присутствия меланопсина, светочувствительного белка. Таким образом, они составляют третий класс фоторецепторов в дополнение к палочек и колбочек.
По сравнению с палочек и колбочек, ipRGC реагируют более медленно и сигнализируют о наличии света в течение длительного времени. Они представляют собой очень небольшую часть (~ 1%) ганглиозных клеток сетчатки. Их функциональные роли не формируют образы и в корне отличаются от функций видения паттернов; они обеспечивают стабильное представление интенсивности окружающего света. У них есть как минимум три основные функции:
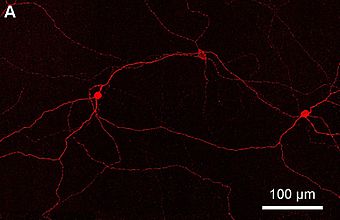 ipRGC, показанный здесь как полное изображение сетчатки от проксимального внутреннего ядерного слоя до слоя ганглиозных клеток с флуоресцентной маркировкой меланопсина
ipRGC, показанный здесь как полное изображение сетчатки от проксимального внутреннего ядерного слоя до слоя ганглиозных клеток с флуоресцентной маркировкой меланопсина Фоторецептивные ганглиозные клетки были выделены у людей, где, помимо регуляции циркадного ритма, они, как было показано, опосредуют определенную степень распознавания света у лиц без стержней и колбочек, страдающих расстройствами палочки и колбочки фоторецепторы. Работа с коллегами показала, что светочувствительные ганглиозные клетки могут иметь некоторую зрительную функцию у людей.
Фотопигмент светочувствительных ганглиозных клеток, меланопсин, возбуждается светом в основном в синей части видимого спектра (пики поглощения ~ 480 нанометров). Механизм фототрансдукции в этих клетках до конца не изучен, но, по-видимому, похож на механизм рабдомерных фоторецепторов беспозвоночных. Помимо прямой реакции на свет, эти клетки могут получать возбуждающее и тормозящее влияние от палочек и колбочек посредством синаптических связей в сетчатке.
Аксоны этих ганглиев иннервируют области мозга, связанные с распознаванием объектов, включая верхний бугорок и дорсальное латеральное коленчатое ядро .
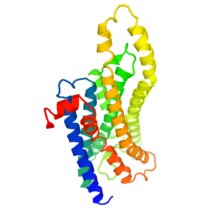 Структура меланопсина
Структура меланопсина Эти фоторецепторные клетки проецируются как через сетчатку, так и в мозг. Они содержат фотопигмент меланопсин в различных количествах вдоль клеточной мембраны, в том числе на аксонах вплоть до диска зрительного нерва, сомы и дендритов клетки. ipRGC содержат мембранные рецепторы нейротрансмиттеров глутамата, глицина и ГАМК. Фоточувствительные ганглиозные клетки реагируют на свет деполяризацией, тем самым увеличивая скорость, с которой они запускают нервные импульсы, что противоположно таковой у других фоторецепторных клеток, которые гиперполяризуются в ответ на свет.
Результаты исследований на мышах предполагают, что аксоны ipRGC являются немиелинизированными.
В отличие от других фоторецепторных пигментов, меланопсин обладает способностью действовать как возбудимый фотопигмент и как фотоизомераза. Вместо того, чтобы требовать, чтобы дополнительные клетки возвращались между двумя изоформами, из полностью транс-ретиналя обратно в 11-цис-ретиналь, прежде чем он сможет подвергнуться другой фототрансдукции, такой как фоторецептор колбочек, которые полагаются на клетки Мюллера и клетки пигментного эпителия сетчатки для этого преобразования, меланопсин способен изомеризовать полностью транс-ретиналь в 11-цис-ретиналь при стимуляции свет без помощи дополнительных ячеек. Две изоформы меланопсина различаются по своей спектральной чувствительности, поскольку изоформа 11-цис-ретиналя более чувствительна к более коротким длинам волн света, в то время как полностью транс-изоформа более чувствительна к более длинным длинам волн света.
 Синаптические входы и выходы ipRGC и их соответствующее расположение в мозге
Синаптические входы и выходы ipRGC и их соответствующее расположение в мозге ipRGC являются как пре-, так и постсинаптическими по отношению к дофаминергическим амакриновым клеткам (DA-клетки) через реципрокные синапсы, при этом ipRGC посылают возбуждающие сигналы DA-клеткам, а DA-клетки посылают тормозные сигналы ipRGC. Эти ингибирующие сигналы передаются через ГАМК, которая совместно высвобождается из DA-клеток вместе с дофамином. Дофамин выполняет функции в процессе адаптации к свету, регулируя транскрипцию меланопсина в ipRGC и тем самым увеличивая чувствительность фоторецепторов. Параллельно с ингибированием амакриновых клеток DA, высвобождающие соматостатин амакриновые клетки, которые сами ингибируются амакриновыми клетками DA, ингибируют ipRGC. Другие синаптические входы для дендритов ipRGC включают биполярные клетки колбочек и биполярные клетки палочек.
Одной постсинаптической мишенью ipRGC является супрахиазматическое ядро (SCN) гипоталамуса, которое служит циркадными часами. в организме. ipRGC высвобождают как белок, активирующий аденилатциклазу гипофиза (PACAP), так и глутамат в SCN через моносинаптическое соединение, называемое ретиногипоталамическим трактом (RHT). Глутамат оказывает возбуждающее действие на нейроны SCN, а PACAP, по-видимому, усиливает эффекты глутамата в гипоталамусе.
Другие постсинаптические мишени ipRGC включают: межжелудочковый листок (IGL), кластер нейронов, расположенный в таламус, который играет роль в циркадном увлечении; оливарное претектальное ядро (OPN), кластер нейронов в среднем мозге, который контролирует зрачковый световой рефлекс; вентролатеральное преоптическое ядро (VLPO), расположенное в гипоталамусе и являющееся центром управления сном; а также миндалины.
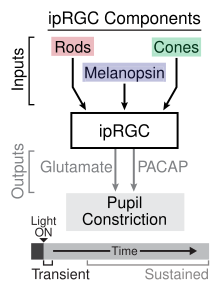 Входы и выходы для ipRGC, участвующих в зрачковом световом рефлексе
Входы и выходы для ipRGC, участвующих в зрачковом световом рефлексе Используя различные мыши с нокаутом фоторецепторов, исследователи идентифицировали роль ipRGC как в кратковременной, так и в постоянной передаче сигналов зрачкового светового рефлекса (PLR). Переходный PLR происходит при яркости от слабого до умеренного и является результатом фототрансдукции, происходящей в стержневых клетках, которые обеспечивают синаптический вход в ipRGC, которые, в свою очередь, передают информацию в оливарное претектальное ядро в среднем мозге. Нейромедиатор, участвующий в передаче информации в средний мозг от ipRGC во временном PLR, - это глутамат. При более ярком свете происходит устойчивый PLR, который включает как фототрансдукцию стержня, обеспечивающего вход в ipRGC, так и фототрансдукцию самих ipRGC через меланопсин. Исследователи предположили, что роль меланопсина в устойчивой PLR связана с его недостаточной адаптацией к световым стимулам в отличие от палочковых клеток, которые демонстрируют адаптацию. Устойчивый PLR поддерживается посредством высвобождения PACAP из ipRGCs в пульсирующем режиме.
Эксперименты с людьми без стержней и колбочек позволили еще одну возможную роль рецептор, который необходимо изучить. В 2007 году для светочувствительной ганглиозной клетки была обнаружена новая роль. Заиди и его коллеги показали, что у людей фоторецептор ганглиозных клеток сетчатки способствует сознательному зрению, а также не формирующим изображения функциям, таким как циркадные ритмы, поведение и зрачковые реакции. Поскольку эти клетки в основном реагируют на синий свет, было высказано предположение, что они играют роль в мезопическом зрении и что старая теория чисто дуплексной сетчатки с стержнем (темным) и колбочкой (свет) световое видение было упрощенным. Таким образом, работа Заиди и его коллег с человеческими объектами без стержней и колбочек также открыла двери для формирования изображений (визуальных) ролей фоторецептора ганглиозных клеток.
Было сделано открытие, что существуют параллельные пути для зрения: один классический стержневой и конусообразный, исходящий из внешней сетчатки, другой - элементарный детектор зрительной яркости, исходящий из внутренней сетчатки. Последнее, кажется, активируется светом раньше первого. Классические фоторецепторы также питаются новой системой фоторецепторов, и постоянство цвета может играть важную роль, как предполагает Фостер.
Авторы модели человека без стержней и колбочек предположили, что рецептор может играть важную роль в понимании многих заболеваний, включая основные причины слепоты во всем мире, такие как глаукома, заболевание, которое влияет на ганглиозные клетки.
У других млекопитающих светочувствительные ганглии играют важную роль в сознательном зрении. Тесты, проведенные Jennifer Ecker et al. обнаружили, что крысы без палочек и колбочек могли научиться плавать к последовательностям вертикальных полос, а не к одинаково люминесцентному серому экрану.
Большинство работ предполагает, что пик спектральная чувствительность рецептора составляет от 460 до 484 нм. Локли и др. в 2003 году показали, что свет с длиной волны 460 нм (синий) подавляет мелатонин вдвое больше, чем свет 555 нм (зеленый), максимальная чувствительность фотопической зрительной системы. В работе Заиди, Локли и соавторов с использованием человека без стержней и колбочек было обнаружено, что очень интенсивный стимул 481 нм привел к некоторому сознательному восприятию света, а это означает, что было реализовано некое рудиментарное зрение.
В 1923 году Клайд Э. Киллер заметил, что зрачки в глазах слепых мышей, которых он случайно вывели, все еще реагируют на свет. Способность мышей без палочек и колбочек сохранять зрачковый световой рефлекс наводила на мысль о дополнительной фоторецепторной клетке.
В 1980-х годах исследования на крысах с дефицитом палочек и колбочек показали регуляцию дофамин в сетчатке, известный нейромодулятор для световой адаптации и фотоэнтренмента.
Исследования продолжались в 1991 году, когда Рассел Г. Фостер и его коллеги, в том числе Игнасио Провенсио, показали что палочки и колбочки не нужны ни для фотоэнтренмента, зрительного привода циркадного ритма, ни для регуляции секреции мелатонина из шишковидной железы через палочку - и мышей-конусов. Более поздняя работа Провенсио и его коллег показала, что этот фотоответ опосредован фотопигментом меланопсином, присутствующим в слое ганглиозных клеток сетчатки.
Фоторецепторы были идентифицированы в 2002 году Самером. Хаттар, Дэвид Берсон и его коллеги, где было показано, что они являются ганглиозными клетками, экспрессирующими меланопсин, которые обладают собственной световой реакцией и проецируются на ряд областей мозга, участвующих в зрении, не формирующем изображение.
В 2005, Панда, Мелян, Цю и его коллеги продемонстрировали, что фотопигмент меланопсина является пигментом фототрансдукции в ганглиозных клетках. Деннис Дэйси и его коллеги показали на одном из видов обезьян Старого Света, что гигантские ганглиозные клетки, экспрессирующие меланопсин, проецируются в латеральное коленчатое ядро (LGN). Ранее были показаны только проекции на средний мозг (претектальное ядро) и гипоталамус (супрахиазматические ядра, SCN). Однако визуальная роль рецептора все еще оставалась неподтвержденной и недоказанной.
Были предприняты попытки выследить рецептор у людей, но люди создавали особые проблемы и требовали новой модели. В отличие от других животных, исследователи не могли этически вызвать потерю палочек и колбочек ни генетически, ни с помощью химических веществ, чтобы непосредственно изучить ганглиозные клетки. В течение многих лет о рецепторе у человека можно было сделать только выводы, хотя временами они были уместными.
В 2007 году Зайди и его коллеги опубликовали свою работу о людях без стержней и конусов, показав, что эти люди сохраняют нормальную реакцию на невизуальные эффекты света. Идентичность фоторецептора без стержня и колбочек у людей была обнаружена в ганглиозных клетках внутренней сетчатки, как было показано ранее на бесстержневых и бесконусных моделях у некоторых других млекопитающих. Работа проводилась на пациентах с редкими заболеваниями, которые приводили к нарушению классической функции фоторецепторов палочки и колбочки, но сохраняли функцию ганглиозных клеток. Несмотря на отсутствие палочек или колбочек, пациенты продолжали демонстрировать циркадный световой поток, циркадные поведенческие паттерны, подавление мелатонина и реакции зрачка с пиковой спектральной чувствительностью к окружающему и экспериментальному свету, которая соответствует фотопигменту меланопсина. Их мозг также мог связывать зрение со светом этой частоты. Клиницисты и ученые сейчас пытаются понять роль нового рецептора в человеческих заболеваниях и слепоте. По своей природе светочувствительные RGC также участвуют в обострении головной боли от света во время приступов мигрени.