
Войти
| Гипоталамус | |
|---|---|
 Расположение гипоталамуса человека Расположение гипоталамуса человека | |
 Расположение гипоталамуса ( синий) по отношению к гипофизу и остальному мозгу Расположение гипоталамуса ( синий) по отношению к гипофизу и остальному мозгу | |
| Подробности | |
| Часть | Головной мозг |
| Идентификаторы | |
| латинский | гипоталамус |
| MeSH | D007031 |
| НейроЛекс ID | birnlex_734 |
| TA98 | A14.1.08.401 A14.1.08.901 |
| TA2 | 5714 |
| FMA | 62008 |
| Анатомические термины нейроанатомии [ редактировать в Викиданных ] | |
Гипоталамуса (от Древнегреческого ὑπό, «под», и θάλαμος, «камера») представляет собой часть мозга, которая содержит ряд небольших ядер с различными функциями. Одна из наиболее важных функций гипоталамуса - связать нервную систему с эндокринной системой через гипофиз. Гипоталамус расположен ниже таламуса и является частью лимбической системы. По терминологии нейроанатомии, он образует вентральную часть промежуточного мозга. Мозг всех позвоночных содержит гипоталамус. У человека он размером с миндаль.
Гипоталамус отвечает за регулирование определенных метаболических процессов и другой деятельности вегетативной нервной системы. Он синтезирует и секретирует определенные нейрогормоны, называемые рилизинг-гормонами или гипоталамическими гормонами, которые, в свою очередь, стимулируют или подавляют секрецию гормонов гипофизом. Гипоталамус контролирует температуру тела, голод, важные аспекты поведения родителей и привязанности, жажду, усталость, сон и циркадные ритмы.
 Гипоталамус человека (показан красным)
Гипоталамус человека (показан красным) Гипоталамус разделен на 3 области (супраоптическая, туберальная, маммиллярная) в парасагиттальной плоскости, что указывает на передне-заднее расположение; и 3 области (перивентрикулярная, медиальная, латеральная) в коронарной плоскости, что указывает на медиально-латеральное расположение. Ядра гипоталамуса расположены в этих конкретных областях и областях. Он обнаружен во всех нервных системах позвоночных. У млекопитающих, магноцеллюлярные нейросекреторные клетки в паравентрикулярном ядре и супраоптическом ядре гипоталамуса производят neurohypophysial гормонов, окситоцин и вазопрессин. Эти гормоны попадают в кровь в задней доле гипофиза. Гораздо меньшие парвоцеллюлярные нейросекреторные клетки, нейроны паравентрикулярного ядра, высвобождают кортикотропин-рилизинг гормон и другие гормоны в портальную систему гипофиза, где эти гормоны диффундируют в переднюю долю гипофиза.
Ядра гипоталамуса включают следующее:
| Область | Площадь | Ядро | Функция |
| Передний (супраоптический) | Преоптический | Преоптическое ядро | |
| Медиальный | Медиальное преоптическое ядро |
| |
| Супраоптическое ядро |
| ||
| Паравентрикулярное ядро |
| ||
| Переднее ядро гипоталамуса |
| ||
| Супрахиазматическое ядро | |||
| Боковой | |||
| Боковое ядро | См. Раздел " Боковой гипоталамус" § Функция - основной источник нейронов орексина, которые проходят через головной и спинной мозг. | ||
| Средний (туберальный) | Медиальный | Дорсомедиальное ядро гипоталамуса |
|
| Вентромедиальное ядро |
| ||
| Дугообразное ядро |
| ||
| Боковой | Боковое ядро | См. Раздел " Боковой гипоталамус" § Функция - основной источник нейронов орексина, которые проходят через головной и спинной мозг. | |
| Боковые туберальные ядра | |||
| Задний (маммиллярный) | Медиальный | Маммиллярные ядра (часть маммиллярных тел ) | |
| Заднее ядро |
| ||
| Боковой | Боковое ядро | См. Раздел " Боковой гипоталамус" § Функция - основной источник нейронов орексина, которые проходят через головной и спинной мозг. | |
| Туберомаммиллярное ядро |
|

Поперечный разрез гипоталамуса обезьяны показывает два основных ядра гипоталамуса по обе стороны от заполненного жидкостью третьего желудочка.

Ядра гипоталамуса

Ядра гипоталамуса на одной стороне гипоталамуса, показанные на трехмерной компьютерной реконструкции.
Гипоталамус тесно связан с другими частями центральной нервной системы, в частности, стволом мозга и его ретикулярной формацией. Как часть лимбической системы, он связан с другими лимбическими структурами, включая миндалину и перегородку, а также связан с областями автономной нервной системы.
Гипоталамус получает много сигналов от ствола мозга, наиболее заметными из которых являются ядро единственного тракта, голубое пятно и вентролатеральный мозг.
Большинство нервных волокон в гипоталамусе проходят двумя путями (двунаправленными).
Некоторые ядра гипоталамуса половой диморфизм ; то есть между мужчинами и женщинами существуют четкие различия как в строении, так и в функциях. Некоторые различия очевидны даже в грубой нейроанатомии: наиболее заметным является половое диморфное ядро в преоптической области, в котором различия заключаются в тонких изменениях в связности и химической чувствительности определенных наборов нейронов. Важность этих изменений можно понять по функциональным различиям между мужчинами и женщинами. Например, самцы большинства видов предпочитают запах и внешний вид самок самцам, что играет важную роль в стимулировании полового поведения самцов. При поражении полового диморфного ядра это предпочтение самок самцами уменьшается. Кроме того, структура секреции гормона роста сексуально диморфна; Вот почему у многих видов взрослых самцов заметно отличить от самок.
Другой поразительный функциональный диморфизм - это поведенческие реакции взрослых на стероиды яичников. Мужчины и женщины реагируют на стероиды яичников по-разному, отчасти потому, что экспрессия чувствительных к эстрогену нейронов в гипоталамусе сексуально диморфна; т.е. рецепторы эстрогена экспрессируются в разных наборах нейронов.
Эстроген и прогестерон могут влиять на экспрессию генов в определенных нейронах или вызывать изменения потенциала клеточной мембраны и активацию киназы, приводя к различным негеномным клеточным функциям. Эстроген и прогестерон связываются со своими родственными рецепторами ядерных гормонов, которые перемещаются в ядро клетки и взаимодействуют с участками ДНК, известными как элементы гормонального ответа (HRE), или привязываются к сайту связывания другого фактора транскрипции. Было показано, что рецептор эстрогена (ER) таким образом трансактивирует другие факторы транскрипции, несмотря на отсутствие элемента ответа на эстроген (ERE) в проксимальной промоторной области гена. В общем, ER и рецепторы прогестерона (PR) являются активаторами генов с увеличением мРНК и последующим синтезом белка после воздействия гормона.
Мужской и женский мозг различаются по распределению рецепторов эстрогена, и это различие является необратимым следствием воздействия неонатальных стероидов. Рецепторы эстрогенов (и рецепторы прогестерона) обнаруживаются в основном в нейронах переднего и медиобазального гипоталамуса, а именно:
 Срединный сагиттальный срез мозга человеческого эмбриона трех месяцев
Срединный сагиттальный срез мозга человеческого эмбриона трех месяцев В неонатальном периоде гонадные стероиды влияют на развитие нейроэндокринного гипоталамуса. Например, они определяют способность женщин проявлять нормальный репродуктивный цикл, а мужчин и женщин - проявлять соответствующее репродуктивное поведение во взрослой жизни.
У приматов влияние андрогенов на развитие менее очевидно, а последствия менее понятны. В головном мозге тестостерон ароматизируется (до эстрадиола ), который является основным активным гормоном, влияющим на развитие. Яички человека секретируют высокий уровень тестостерона примерно с 8-й недели жизни плода до 5-6 месяцев после рождения (аналогичный перинатальный всплеск тестостерона наблюдается у многих видов), процесс, который, по-видимому, лежит в основе мужского фенотипа. Эстроген из материнского кровотока относительно неэффективен, отчасти из-за высокого уровня циркулирующих стероидсвязывающих белков во время беременности.
Половые стероиды - не единственные важные факторы, влияющие на развитие гипоталамуса; в частности, предпубертатный стресс в раннем возрасте (у крыс) определяет способность взрослого гипоталамуса реагировать на острый стрессор. В отличие от рецепторов гонадных стероидов, рецепторы глюкокортикоидов очень широко распространены по всему мозгу; в паравентрикулярном ядре они опосредуют контроль синтеза и секреции CRF с отрицательной обратной связью, но в других местах их роль недостаточно изучена.
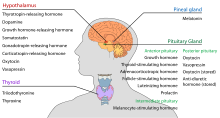 Эндокринные железы головы и шеи человека и их гормоны
Эндокринные железы головы и шеи человека и их гормоны Гипоталамус выполняет центральную нейроэндокринную функцию, в первую очередь за счет контроля над передней долей гипофиза, которая, в свою очередь, регулирует различные эндокринные железы и органы. Релизинг-гормоны (также называемые рилизинг-факторами) продуцируются в ядрах гипоталамуса, а затем транспортируются по аксонам либо к срединному возвышению, либо к задней доле гипофиза, где они сохраняются и высвобождаются по мере необходимости.
В гипоталамо-аденогипофизарной оси рилизинг-гормоны, также известные как гипофизиотропные или гипоталамические гормоны, высвобождаются из среднего возвышения, удлинения гипоталамуса, в портальную систему гипофиза, которая переносит их в переднюю долю гипофиза, где они выполняют свои регулирующие функции. на секрецию гормонов аденогипофиза. Эти гипофизиотропные гормоны стимулируются парвоцеллюлярными нейросекреторными клетками, расположенными в перивентрикулярной области гипоталамуса. После попадания в капилляры третьего желудочка гипофизиотропные гормоны проходят через так называемый гипоталамо-гипофизарный портальный кровоток. Достигнув места назначения в передней доле гипофиза, эти гормоны связываются со специфическими рецепторами, расположенными на поверхности клеток гипофиза. В зависимости от того, какие клетки активируются посредством этого связывания, гипофиз либо начнет секретировать, либо прекратит выделять гормоны в остальную часть кровотока.
Другие гормоны, выделяемые средним возвышением, включают вазопрессин, окситоцин и нейротензин.
В гипоталамо-нейрогипофизарной оси нейрогипофизарные гормоны выделяются в кровоток из заднего гипофиза, который на самом деле является продолжением гипоталамуса.
| Секретируемый гормон | Сокращенное название | Произведено | Эффект |
|---|---|---|---|
| Окситоцин | OXY или OXT | Магноцеллюлярные нейросекреторные клетки паравентрикулярного ядра и супраоптического ядра | Сокращение матки Лактация (рефлекс расслабления) |
| Вазопрессин (антидиуретический гормон) | ADH или AVP | Магноцеллюлярные и парвоцеллюлярные нейросекреторные клетки паравентрикулярного ядра, магноцеллюлярные клетки супраоптического ядра | Повышение проницаемости для воды клеток дистального канальца и собирательного канала в почках, что способствует реабсорбции воды и выведению концентрированной мочи. |
Также известно, что гормоны гипоталамо-гипофизарно-надпочечниковой системы (HPA) связаны с некоторыми кожными заболеваниями и гомеостазом кожи. Существуют доказательства связи гиперактивности гормонов HPA со стрессовыми кожными заболеваниями и кожными опухолями.
Гипоталамус координирует многие гормональные и поведенческие циркадные ритмы, сложные паттерны нейроэндокринных процессов, сложные гомеостатические механизмы и важные формы поведения. Следовательно, гипоталамус должен реагировать на множество различных сигналов, некоторые из которых генерируются извне, а некоторые - внутренне. Передача сигналов дельта-волной, возникающая либо в таламусе, либо в коре головного мозга, влияет на секрецию высвобождающих гормонов; GHRH и пролактин стимулируются, в то время как TRH ингибируется.
Гипоталамус реагирует на:
Обонятельные стимулы важны для полового размножения и нейроэндокринной функции у многих видов. Например, если беременная мышь подвергается воздействию мочи «незнакомого» самца в критический период после полового акта, беременность не наступает ( эффект Брюса ). Таким образом, во время полового акта у самки мыши формируется точная «обонятельная память» о своем партнере, которая сохраняется в течение нескольких дней. Феромональные сигналы помогают синхронизировать эструс у многих видов; у женщин синхронизированная менструация может также возникать из-за феромонных сигналов, хотя роль феромонов у людей оспаривается.
Пептидные гормоны оказывают важное влияние на гипоталамус, и для этого они должны проходить через гематоэнцефалический барьер. Гипоталамус частично ограничен специализированными областями мозга, в которых отсутствует эффективный гематоэнцефалический барьер; капиллярный эндотелий на этих сайтах Фенестрированный, чтобы позволить свободное прохождение даже больших белков и других молекул. Некоторые из этих участков являются участками нейросекреции - нейрогипофиз и срединное возвышение. Однако в других местах мозг анализирует состав крови. Два из этих сайтов, СФО ( subfornical органа ) и OVLT ( Organum vasculosum из пластинки терминального ) являются так называемыми circumventricular органов, где нейроны находятся в тесном контакте как с кровью и CSF. Эти структуры плотно васкуляризированы и содержат осморецептивные и натрий-рецептивные нейроны, которые контролируют питье, высвобождение вазопрессина, экскрецию натрия и аппетит натрия. Они также содержат нейроны с рецепторами ангиотензина, предсердного натрийуретического фактора, эндотелина и релаксина, каждый из которых важен для регуляции баланса жидкости и электролитов. Нейроны в OVLT и SFO проецируются в супраоптическое ядро и паравентрикулярное ядро, а также в преоптические области гипоталамуса. Окружные желудочковые органы также могут быть местом действия интерлейкинов, вызывающих как лихорадку, так и секрецию АКТГ посредством воздействия на паравентрикулярные нейроны.
Неясно, как все пептиды, влияющие на гипоталамическую активность, получают необходимый доступ. В случае пролактина и лептина есть данные об их активном поглощении сосудистым сплетением из крови в спинномозговую жидкость (CSF). Некоторые гормоны гипофиза оказывают отрицательное влияние на секрецию гипоталамуса; например, гормон роста действует на гипоталамус, но неясно, как он попадает в мозг. Есть также данные о центральном действии пролактина.
Полученные данные свидетельствуют о том, что гормон щитовидной железы (Т4) поглощается глиальными клетками гипоталамуса в инфундибулярном ядре / срединном возвышении, и что здесь он превращается в Т3 дейодиназой 2 типа (D2). После этого Т3 транспортируется в нейроны, продуцирующие тиреотропин-рилизинг-гормон ( TRH ), в паравентрикулярном ядре. В этих нейронах были обнаружены рецепторы гормонов щитовидной железы, что указывает на их чувствительность к стимулам Т3. Кроме того, эти нейроны экспрессировали MCT8, переносчик гормонов щитовидной железы, что подтверждает теорию о том, что Т3 транспортируется в них. Затем Т3 может связываться с рецептором гормона щитовидной железы в этих нейронах и влиять на выработку тиреотропин-рилизинг-гормона, тем самым регулируя выработку гормона щитовидной железы.
Гипоталамус функционирует как своего рода термостат для тела. Он устанавливает желаемую температуру тела и стимулирует выработку и удержание тепла для повышения температуры крови до более высокого уровня или потоотделение и расширение сосудов для охлаждения крови до более низкой температуры. Все лихорадки возникают из-за повышенного давления в гипоталамусе; повышенная температура тела по любой другой причине классифицируется как гипертермия. В редких случаях прямое повреждение гипоталамуса, например, в результате инсульта, вызывает повышение температуры тела; это иногда называют гипоталамической лихорадкой. Однако чаще такое повреждение вызывает аномально низкую температуру тела.
Гипоталамус содержит нейроны, которые сильно реагируют на стероиды и глюкокортикоиды - стероидные гормоны надпочечников, высвобождаемые в ответ на АКТГ. Он также содержит специализированные чувствительные к глюкозе нейроны (в дугообразном ядре и вентромедиальном гипоталамусе ), которые важны для аппетита. В преоптической области располагаются термочувствительные нейроны; они важны для секреции TRH.
Секреция окситоцина в ответ на сосание или вагино-цервикальную стимуляцию опосредуется некоторыми из этих путей; Секреция вазопрессина в ответ на сердечно-сосудистые стимулы, исходящие от хеморецепторов в теле сонной артерии и дуги аорты, а также от рецепторов предсердного объема низкого давления, опосредуется другими. У крыс стимуляция влагалища также вызывает секрецию пролактина, что приводит к ложной беременности после бесплодного спаривания. У кролика половой акт вызывает рефлекторную овуляцию. У овец стимуляция шейки матки в присутствии высоких уровней эстрогена может вызвать материнское поведение у девственной овцы. Все эти эффекты опосредуются гипоталамусом, и информация передается в основном по спинным путям, которые передаются в ствол мозга. Стимуляция сосков стимулирует высвобождение окситоцина и пролактина и подавляет высвобождение ЛГ и ФСГ.
Сердечно-сосудистые стимулы переносятся блуждающим нервом. Блуждающий нерв также передает разнообразную висцеральную информацию, включая, например, сигналы, возникающие при растяжении или опорожнении желудка, для подавления или стимулирования кормления, сигнализируя о высвобождении лептина или гастрина, соответственно. И снова эта информация достигает гипоталамуса через реле в стволе мозга.
Кроме того, функция гипоталамуса реагирует и регулируется уровнями всех трех классических нейромедиаторов моноаминов, норадреналина, дофамина и серотонина (5-гидрокситриптамина) в тех трактах, из которых он получает иннервацию. Например, норадренергические входы, возникающие из голубого пятна, имеют важные регулирующие эффекты на уровни кортикотропин-рилизинг-гормона (CRH).
| Пептиды, улучшающие пищевое поведение | Пептиды, снижающие пищевое поведение |
|---|---|
| Грелин | Лептин |
| Нейропептид Y | (α, β, γ) - гормоны, стимулирующие меланоциты |
| Агути-родственный пептид | Пептиды транскрипции, регулируемые кокаином и амфетамином |
| Орексины (А, В) | Кортикотропин-рилизинг-гормон |
| Меланин-концентрирующий гормон | Холецистокинин |
| Галанин | Инсулин |
| Глюкагоноподобный пептид 1 |
Крайняя боковая часть вентромедиального ядра гипоталамуса отвечает за контроль пищевого потребления. Стимуляция этой области вызывает повышенное потребление пищи. Двустороннее поражение этой области вызывает полное прекращение приема пищи. Медиальные части ядра оказывают контролирующее влияние на латеральную часть. Двустороннее поражение медиальной части вентромедиального ядра вызывает гиперфагию и ожирение животного. Дальнейшее поражение боковой части вентромедиального ядра у того же животного приводит к полному прекращению приема пищи.
Существуют разные гипотезы, связанные с этим регулированием:
Медиальная зона гипоталамуса является частью цепи, которая контролирует мотивированное поведение, например, защитное поведение. Анализ Fos- метки показал, что ряд ядер в «столбце поведенческого контроля» важен для регуляции выражения врожденного и условного защитного поведения.
Контакт с хищником (например, с кошкой) вызывает у лабораторных грызунов защитное поведение, даже если животное никогда не контактировало с кошкой. В гипоталамусе это воздействие вызывает увеличение количества Fos-меченных клеток в переднем ядре гипоталамуса, дорсомедиальной части вентромедиального ядра и в вентролатеральной части премамиллярного ядра (PMDvl). Премамиллярное ядро играет важную роль в выражении защитного поведения по отношению к хищнику, поскольку повреждения в этом ядре отменяют защитное поведение, такое как замирание и бегство. PMD не модулирует защитное поведение в других ситуациях, поскольку поражения этого ядра оказали минимальное влияние на баллы замораживания после шока. PMD имеет важные связи с дорсальной периакведуктальной серой, важной структурой в выражении страха. Кроме того, животные демонстрируют поведение оценки риска для окружающей среды, ранее связанной с кошкой. Анализ клеток, меченных Fos, показал, что PMDv1 является наиболее активированной структурой в гипоталамусе, а инактивация мусцимолом перед воздействием контекста отменяет защитное поведение. Следовательно, гипоталамус, в основном PMDv1, играет важную роль в выражении врожденного и условного защитного поведения хищника.
Точно так же гипоталамус играет роль в социальном поражении : ядра в медиальной зоне также мобилизуются во время встречи с агрессивным сородичем. У побежденного животного наблюдается повышение уровней Fos в половых диморфных структурах, таких как медиальное предоптическое ядро, вентролатеральная часть вентромедиального ядра и вентральное премаммилярное ядро. Такие структуры важны в других формах социального поведения, таких как сексуальное и агрессивное поведение. Более того, мобилизируется и премамиллярное ядро, дорсомедиальная часть, но не вентролатеральная часть. Поражения в этом ядре отменяют пассивное защитное поведение, такое как замораживание и поза «на спине».


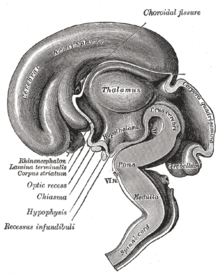
Человеческий мозг слева в разрезе срединно-сагиттальный вид

Расположение гипоталамуса