
Войти
| Аденоассоциированный вирус | |
|---|---|
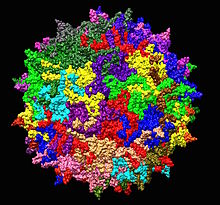 | |
| Структура аденоассоциированного вируса серотипа 2 из 1LP3. В центре показана одна пятеричная ось. | |
| Научная классификация | |
| (без рейтинга): | Вирус |
| Царство: | Моноднавирия |
| Королевство: | Шотокувира |
| Тип: | Cossaviricota |
| Класс: | Quintoviricetes |
| Порядок: | Piccovirales |
| Семейство: | Parvoviridae |
| Подсемейство: | Parvovirinae |
| Род: | Dependoparvovirus |
| Включены вирусы: | |
| |
аденоассоциированные вирусы (AAV ) - небольшие вирусы, которые инфицируют людей и некоторые другие виды приматов. Они принадлежат к роду Dependoparvovirus, который, в свою очередь, принадлежит к семейству Parvoviridae. Они небольшие (20 nm ) репликационно-дефектных, вирусов без оболочки.
AAV, как сейчас известно, вызывают заболевание. Вирусы вызывают очень слабый иммунный ответ. Несколько дополнительных функций делают AAV привлекательным кандидатом для создания вирусных векторов для генной терапии и для создания моделей изогенных заболеваний человека. Генная терапия векторами с использованием AAV может инфицировать как делящиеся, так и покоящиеся клетки и сохраняться во внехромосомном состоянии, не интегрируясь в геном клетки-хозяина, хотя при интеграции нативного вируса Вирусно переносимые гены в геном хозяина действительно происходят. Интеграция может быть важна для определенных приложений, но также может иметь нежелательные последствия. Недавние клинические испытания на людях с использованием AAV для генной терапии сетчатки показали многообещающие результаты.
Аденоассоциированный вирус (AAV), который ранее считался контаминантом в препаратах аденовируса, был впервые идентифицирован как зависимый парвовирус в 1960-х годах в лабораториях Боба Атчисона в Питтсбурге и Уоллеса Роу в NIH. Серологические исследования на людях впоследствии показали, что, несмотря на присутствие у людей, инфицированных , такими как аденовирус или вирус герпеса, сам по себе AAV не вызывает каких-либо заболеваний.
AAV дикого типа вызвали значительный интерес исследователей генной терапии из-за ряда особенностей. Главным из них является очевидное отсутствие патогенности вируса. Он также может инфицировать неделящиеся клетки и обладает способностью стабильно интегрироваться в геном клетки-хозяина в конкретном сайте (обозначенном AAVS1) в хромосоме 19 человека. Эта особенность делает его несколько более предсказуемым, чем ретровирусы, которые представляют угрозу случайного внедрения и мутагенеза, за которым иногда следует развитие рака. Геном AAV наиболее часто интегрируется в упомянутый сайт, тогда как случайные включения в геном происходят с незначительной частотой. Однако разработка AAV в качестве векторов для генной терапии устранила эту интегративную способность путем удаления rep и кэпа из ДНК вектора. Желаемый ген вместе с промотором для управления транскрипцией гена вставляется между инвертированными концевыми повторами (ITR), которые способствуют образованию конкатемера в ядре после одноцепочечной векторной ДНК. превращается комплексами ДНК-полимеразы клетки-хозяина в двухцепочечную ДНК. Векторы генной терапии на основе AAV образуют эписомальные конкатемеры в ядре клетки-хозяина. В неделящихся клетках эти конкатемеры остаются нетронутыми в течение всей жизни клетки-хозяина. В делящихся клетках ДНК AAV теряется в результате деления клеток, поскольку эписомальная ДНК не реплицируется вместе с ДНК клетки-хозяина. Случайная интеграция ДНК AAV в геном хозяина обнаруживается, но происходит с очень низкой частотой. AAV также обладают очень низкой иммуногенностью, по-видимому, ограниченной образованием нейтрализующих антител, в то время как они не вызывают четко выраженного цитотоксического ответа. Эта особенность, наряду со способностью инфицировать покоящиеся клетки, свидетельствует об их преобладании над аденовирусами как векторами для генной терапии.
человека. Использование вируса действительно имеет некоторые недостатки. Способность клонирования вектора относительно ограничена, и для большинства терапевтических генов требуется полная замена 4,8-килобазного генома вируса. Поэтому большие гены не подходят для использования в стандартном векторе AAV. В настоящее время изучаются варианты преодоления ограниченных возможностей кодирования. ITR AAV двух геномов могут отжигаться с образованием конкатемеров «голова к хвосту», что почти удваивает емкость вектора. Вставка сайтов сплайсинга позволяет удалить ITR из транскрипта.
Благодаря преимуществам специализированной генной терапии AAV, исследователи создали измененную версию AAV, названную самокомплементарным аденоассоциированным вирусом (scAAV). В то время как AAV упаковывает одну цепь ДНК и должен ждать, пока ее вторая цепь будет синтезирована, scAAV упаковывает две более короткие цепи, которые комплементарны друг другу. Избегая синтеза второй цепи, scAAV может экспрессироваться быстрее, хотя, как предостережение, scAAV может кодировать только половину и без того ограниченной емкости AAV. Недавние сообщения предполагают, что векторы scAAV более иммуногенны, чем одноцепочечные аденовирусные векторы, вызывая более сильную активацию цитотоксических Т-лимфоцитов.
Считается, что гуморальный иммунитет, вызванный инфекцией дикого типа, является обычным явлением. Связанная с этим нейтрализующая активность ограничивает применимость наиболее часто используемого серотипа AAV2 в определенных приложениях. Соответственно, большинство текущих клинических испытаний включает доставку AAV2 в мозг, относительно иммунологически привилегированный орган. В головном мозге AAV2 сильно нейрон-специфичен.
На сегодняшний день векторы AAV были использованы в более чем 250 клинических испытаниях по всему миру, что составляет примерно 8,3% испытаний генной терапии с вирусным вектором. Недавно были получены многообещающие результаты испытаний фазы 1 и фазы 2 для ряда заболеваний, включая врожденный амавроз Лебера, гемофилию, застойную сердечную недостаточность, спинальная мышечная атрофия, дефицит липопротеинлипазы и болезнь Паркинсона.
| Показание | Ген | Способ введения | Фаза | Номер субъекта | Статус |
| Муковисцидоз | CFTR | Легкое, аэрозоль | I | 12 | Завершено |
| CFTR | Легкое, через аэрозоль | II | 38 | Полное | |
| CFTR | Легкое, через аэрозоль | II | 100 | Полное | |
| Гемофилия B | FIX | Внутримышечный | I | 9 | Полный |
| FIX | Печеночная артерия | I | 6 | Закончился | |
| Артрит | TNFR: Fc | Внутрисуставной | I | 1 | Продолжается |
| Наследственная эмфизема | ААТ | Внутримышечный | I | 12 | Текущий |
| Врожденный амавроз Лебера | RPE65 | Субретинальный | I – II | Множественный | Несколько текущих и полное |
| Возрастная дегенерация желтого пятна | sFlt-1 | Субретинальная | I – II | 24 | Постоянно |
| Мышечная дистрофия Дюшенна | SGCA | Внутримышечно | I | 10 | Текущая |
| Болезнь Паркинсона | GAD65, GAD67 | Внутричерепная | I | 12 | Полная |
| Болезнь Канавана | Внутричерепная | I | 21 | Текущая | |
| Болезнь Баттена | CLN2 | Внутричерепное | I | 10 | Постоянно |
| Болезнь Альцгеймера | NGF | Внутричерепное | I | 6 | Постоянно |
| Спинальная мышечная атрофия | SMN1 | Внутривенно и интратекально | I – III | 15 | Несколько текущих и полных |
| Застойная сердечная недостаточность | Внутрикоронарная | IIb | 250 | Постоянно |
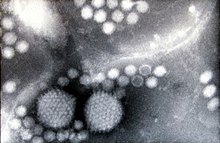 Две частицы аденовируса, окруженные многочисленные аденоассоциированные вирусы меньшего размера (электронная микроскопия с негативным окрашиванием, увеличение примерно 200000 ×)
Две частицы аденовируса, окруженные многочисленные аденоассоциированные вирусы меньшего размера (электронная микроскопия с негативным окрашиванием, увеличение примерно 200000 ×) Геном AAV построен из одноцепочечной дезоксирибонуклеиновой кислоты (ss ДНК ), либо положительно, либо отрицательно, что примерно 4,7 килобазы в длину. Геном содержит ITR на обоих концах цепи ДНК и две открытые рамки считывания (ORF): rep и cap. Первый состоит из четырех перекрывающихся генов, кодирующих Rep-белки, необходимые для жизненного цикла AAV, а последний содержит перекрывающиеся нуклеотидные последовательности белков капсида : VP1, VP2 и VP3, которые взаимодействуют с образуют капсид с икосаэдрической симметрией.
Последовательности инвертированных концевых повторов (ITR) содержат 145 оснований каждая. Они были названы так из-за их симметрии, которая, как было показано, необходима для эффективного размножения генома AAV. Особенностью этих последовательностей, которая придает им это свойство, является их способность образовывать шпильку, которая способствует так называемому самопраймированию, которое позволяет примазе -независимый синтез второй цепи ДНК.. Было также показано, что ITR необходимы как для интеграции ДНК AAV в геном клетки-хозяина (19-я хромосома у человека), так и для выхода из него, а также для эффективного инкапсидации ДНК AAV в сочетании с генерацией полностью собранных, устойчивых к дезоксирибонуклеазе частиц AAV.
Что касается генной терапии, ITR кажутся единственными последовательностями, необходимыми в цис рядом с терапевтическим геном : структурные (кэп) и упаковочные (реп) белки могут доставляться в транс. Исходя из этого предположения, было разработано множество способов эффективного получения рекомбинантных векторов AAV (rAAV), содержащих репортер или терапевтический ген. Однако было также опубликовано, что ITR - не единственные элементы, необходимые в странах СНГ для эффективной репликации и инкапсидации. Несколько исследовательских групп идентифицировали последовательность, обозначенную цис-действующим Rep-зависимым элементом (CARE) внутри кодирующей последовательности реп-гена. Было показано, что CARE усиливает репликацию и инкапсидацию, когда присутствует в цис.
На «левой стороне» генома есть два промотора называемые p5 и p19, из которых могут быть получены две перекрывающиеся матричные рибонуклеиновые кислоты (мРНК ) разной длины. Каждый из них содержит интрон, который может быть либо сплайсирован, либо нет. Учитывая эти возможности, можно синтезировать четыре различных мРНК и, следовательно, четыре различных белка Rep с перекрывающимися последовательностями. Их имена обозначают их размеры в килодальтон (кДа): Rep78, Rep68, Rep52 и Rep40. Rep78 и 68 могут специфически связывать шпильку, образованную ITR, в акте самопраймирования и расщеплять в конкретной области, обозначенной конечным сайтом разрешения, внутри шпильки. Также было показано, что они необходимы для AAVS1-специфической интеграции генома AAV. Было показано, что все четыре Rep-белка связывают АТФ и обладают активностью геликазы. Также было показано, что они активируют транскрипцию с промотора p40 (упомянутого ниже), но подавляют как промоторы p5, так и p19.
Правая сторона положительного - воспринимаемый геном AAV кодирует перекрывающиеся последовательности трех капсидных белков, VP1, VP2 и VP3, которые начинаются с одного промотора, обозначенного p40. Молекулярные массы этих белков составляют 87, 72 и 62 килодальтона соответственно. Капсид AAV состоит из смеси VP1, VP2 и VP3, всего 60 мономеров, расположенных с симметрией икосаэдра в соотношении 1: 1: 10, с расчетным размером 3,9 мегадальтона. Кристаллическая структура белка VP3 была определена Xie, Bue и др.
 капсидом AAV2, показанным в виде ленточной диаграммы, задняя половина которого скрыта для ясности. В центре показана одна пятая ось симметрии.
капсидом AAV2, показанным в виде ленточной диаграммы, задняя половина которого скрыта для ясности. В центре показана одна пятая ось симметрии. Ген cap продуцирует дополнительный неструктурный белок, называемый белком, активирующим сборку (AAP). Этот белок вырабатывается из ORF2 и необходим для процесса сборки капсида. Точная функция этого белка в процессе сборки и его структура до сих пор не решены.
Все три VP транслируются с одной мРНК. После того, как эта мРНК синтезирована, она может быть сплайсирована двумя разными способами: более длинный или более короткий интрон может быть вырезан, что приведет к образованию двух пулов мРНК: 2,3 т.п.н. и пул мРНК длиной 2,6 т.п.н. Обычно, особенно в присутствии аденовируса, более длинный интрон является предпочтительным, поэтому мРНК длиной 2,3 т.п.н. представляет собой так называемый «основной сплайсинг». В этой форме первый кодон AUG, с которого начинается синтез белка VP1, вырезан, что приводит к снижению общего уровня синтеза белка VP1. Первый кодон AUG, который остается в основном сплайсинге, является кодоном инициации для белка VP3. Однако выше этого кодона в той же открытой рамке считывания лежит последовательность ACG (кодирующая треонин), которая окружена оптимальным контекстом Козака. Это способствует низкому уровню синтеза белка VP2, который на самом деле является белком VP3 с дополнительными N-концевыми остатками, как и VP1.
Так как более крупный интрон предпочтительнее для сплайсинга, и поскольку в основном сплайсинге кодон ACG представляет собой гораздо более слабый сигнал инициации трансляции , соотношение, при котором структурные белки AAV синтезируются in vivo, составляет примерно 1: 1: 20, что является таким же, как и в зрелой вирусной частице. Было показано, что уникальный фрагмент на N-конце белка VP1 обладает активностью фосфолипазы A2 (PLA2), которая, вероятно, необходима для высвобождения частиц AAV из поздних эндосом. Muralidhar et al. сообщили, что VP2 и VP3 имеют решающее значение для правильной сборки вириона. Однако совсем недавно Warrington et al. показал, что VP2 не является необходимым для полного образования вирусных частиц и эффективной инфекционности, а также представил, что VP2 может выдерживать большие вставки на своем N-конце, в то время как VP1 не может, вероятно, из-за присутствия домена PLA2.
Два вида AAV были признаны Международным комитетом по таксономии вирусов в 2013 г.: аденоассоциированный зависимый парвовирус A (ранее AAV-1, -2, -3 и -4) и аденоассоциированный зависимый парвовирус B (ранее AAV-5).
До 1990-х годов практически вся биология AAV изучалась с использованием AAV серотипа 2. Однако AAV широко распространен у людей и других приматы и несколько серотипов были выделены из различных образцов тканей. Серотипы 2, 3, 5 и 6 были обнаружены в клетках человека, серотипы AAV 1, 4 и 7–11 - в образцах нечеловеческих приматов. По состоянию на 2006 год было описано 11 серотипов AAV , 11-й - в 2004 году. Капсидные белки AAV содержат 12 гипервариабельных участков поверхности, при этом наибольшая вариабельность проявляется в трехкратных проксимальных пиках, но геном парвовируса в целом является высококонсервативным. репликационные и структурные гены по серотипам. Все известные серотипы могут инфицировать клетки из множества различных типов тканей. Тканевая специфичность определяется серотипом капсида, и псевдотипирование векторов AAV, чтобы изменить диапазон их тропизма, вероятно, будет важным для их использования в терапии.
Серотип 2 (AAV2) до сих пор изучался наиболее широко. AAV2 представляет собой естественный тропизм к скелетным мышцам, нейронам, клеткам гладкой мускулатуры сосудов и гепатоцитам.
Для AAV2 были описаны три клеточных рецептора: гепарансульфат протеогликан (HSPG), Vβ5интегрин и рецептор 1 фактора роста фибробластов (FGFR-1). Первый функционирует как первичный рецептор, в то время как последние два обладают корецепторной активностью и позволяют AAV проникать в клетку посредством рецептор-опосредованного эндоцитоза. Эти результаты исследования были оспорены Qiu, Handa, et al. HSPG функционирует как первичный рецептор, хотя его изобилие во внеклеточном матриксе может улавливать частицы AAV и снижать эффективность инфекции.
Исследования показали, что серотип 2 вируса (AAV-2) очевидно, убивает раковые клетки, не нанося вреда здоровым. «Наши результаты показывают, что аденоассоциированный вирус типа 2, который инфицирует большую часть населения, но не имеет известных побочных эффектов, убивает несколько типов раковых клеток, но не влияет на здоровые клетки», - сказал Крейг Мейерс, профессор иммунология и микробиология в медицинском колледже Penn State в Пенсильвании в 2005 году. Это могло привести к созданию нового противоракового агента.
Хотя AAV2 является наиболее популярным серотипом в различных исследованиях, основанных на AAV, было показано, что другие серотипы могут быть более эффективными в качестве векторов доставки генов. Например, AAV6 гораздо лучше поражает эпителиальные клетки дыхательных путей, AAV7 демонстрирует очень высокую скорость трансдукции мышечных клеток скелетных мышц (аналогично AAV1 и AAV5), AAV8 превосходно трансдуцирует гепатоциты, а AAV1 и 5 оказались очень эффективными в доставке генов. к эндотелиальным клеткам сосудов. В головном мозге большинство серотипов AAV проявляют нейрональный тропизм, в то время как AAV5 также трансдуцирует астроциты. AAV6, гибрид AAV1 и AAV2, также проявляет более низкую иммуногенность, чем AAV2.
Серотипы могут различаться в отношении рецепторов, с которыми они связаны. Например, трансдукция AAV4 и AAV5 может ингибироваться растворимыми сиаловыми кислотами (в различной форме для каждого из этих серотипов), и было показано, что AAV5 проникает в клетки через тромбоцитарный фактор роста рецептор.
Было предпринято много усилий для создания и улучшения новых вариантов AAV как для клинических, так и для исследовательских целей. Такие модификации включают новые тропизмы, направленные на конкретные ткани, и модифицированные поверхностные остатки, чтобы избежать обнаружения иммунной системой. Помимо выбора конкретных штаммов рекомбинантного AAV (rAAV) для нацеливания на определенные клетки, исследователи также исследовали псевдотипирование AAV, практику создания гибридов определенных штаммов AAV для достижения еще более точной цели. Гибрид создается путем взятия капсида из одного штамма и генома из другого штамма. Например, исследование с участием AAV2 / 5, гибрида с геномом AAV2 и капсидом AAV5, позволило достичь большей точности и диапазона в клетках мозга, чем AAV2, которого удалось бы достичь без гибридизации. Исследователи продолжили эксперименты с псевдотипированием, создав штаммы с гибридными капсидами. AAV-DJ имеет гибридный капсид из восьми различных штаммов AAV; как таковой, он может инфицировать различные клетки во многих областях тела - свойство, которым не обладает один штамм AAV с ограниченным тропизмом. Другие усилия по разработке и улучшению новых вариантов AAV включали реконструкцию предков вирусов для создания новых векторов с улучшенными свойствами для клинического применения и изучение биологии AAV.
AAV имеет особый интерес для генных терапевтов из-за его очевидной ограниченной способности вызывать иммунные ответы у людей, фактор, который должен положительно влиять на эффективность векторной трансдукции при одновременном снижении риска любой иммуно-ассоциированной патологии.
AAV не считается имеющим никаких известная роль в заболевании.
врожденный иммунный ответ на векторы AAV был охарактеризован на животных моделях. Внутривенное введение мышам вызывает кратковременную продукцию провоспалительных цитокинов и некоторую инфильтрацию нейтрофилов и других лейкоцитов в печень, что, по-видимому, секвестрирует большую часть введенные вирусные частицы. Как уровни растворимого фактора, так и клеточная инфильтрация возвращаются к исходному уровню в течение шести часов. Напротив, более агрессивные вирусы вызывают врожденные реакции, длящиеся 24 часа или дольше.
Известно, что вирус вызывает устойчивый гуморальный иммунитет на животных моделях и у людей. популяция, где до 80% людей считаются серопозитивными в отношении AAV2. Антитела, как известно, нейтрализуют, и для применения в генной терапии они действительно влияют на эффективность векторной трансдукции через некоторые пути введения. Наряду с постоянными уровнями специфических антител к AAV, как показывают исследования с первичной бустом на животных, так и клинические испытания, B-клеточная память также сильна. У серопозитивных людей циркулирующие IgG антитела к AAV2, по-видимому, в основном состоят из подклассов IgG1 и IgG2, с небольшим количеством или отсутствием IgG3 или IgG4.
Цикл заражения AAV состоит из нескольких этапов, от инфицирования клетки до производства новых инфекционных частиц:
Некоторые из этих шагов могут выглядеть по-разному в разных типах клеток, что отчасти способствует defi Нед и довольно ограниченный природный тропизм AAV. Репликация вируса также может варьироваться для одного типа клеток, в зависимости от текущей фазы клеточного цикла клетки.
Характерной чертой аденоассоциированного вируса является недостаточность репликации и, следовательно, его невозможность размножаться в непораженных клетках. Аденоассоциированный вирус распространяется путем совместного инфицирования клетки вспомогательным вирусом. Первым вспомогательным вирусом, который был описан как обеспечивающий успешную генерацию новых частиц AAV, был аденовирус, от которого произошло название AAV. Затем было показано, что репликации AAV могут способствовать выбранные белки, происходящие из генома аденовируса, другие вирусы, такие как HSV или коровья оспа, или генотоксические агенты, такие как УФ облучение или гидроксимочевина <163.>. В зависимости от наличия или отсутствия вируса-помощника жизненный цикл AAV следует либо по литическому, либо по лизогенному пути, соответственно. Если есть вирус-помощник, экспрессия гена AAV активируется, позволяя вирусу реплицироваться с использованием полимеразы клетки-хозяина. Когда вирус-помощник убивает клетку-хозяина, высвобождаются новые вирионы AAV. Если вирус-помощник отсутствует, AAV проявляет лизогенное поведение. Когда AAV заражает только клетку, экспрессия его гена подавляется (AAV не реплицируется), и его геном включается в геном хозяина (в хромосому 19 человека). В редких случаях лизис может происходить без вспомогательного вируса, но обычно AAV не может реплицироваться и убивать клетку сам по себе.
Минимальный набор аденовирусных генов, необходимый для эффективного генерирования дочерних частиц AAV, был обнаружен Мацуситой, Эллингером и др. Это открытие позволило разработать новые методы получения рекомбинантных AAV, которые не требуют совместной инфекции аденовирусами продуцирующих AAV клеток. В отсутствие вируса-помощника или генотоксических факторов ДНК AAV может либо интегрироваться в геном хозяина, либо сохраняться в эписомальной форме. В первом случае интеграция опосредуется белками Rep78 и Rep68 и требует наличия ITR, фланкирующих интегрируемую область. У мышей геном AAV сохранялся в течение длительных периодов времени в покоящихся тканях, таких как скелетные мышцы, в эписомальной форме (круглая конформация голова к хвосту).